Реферат: Размножение. Доклад: Бесполое и половое размножение
Как правило, в размножении животных участвуют две особи — самец и самка. Для этого у них имеются половые органы . К мужским половым органам животных относятся семенники , а к женским — яичники . В них образуются половые клетки — соответственно сперматозоиды и яйцеклетки .
Чтобы образовался новый организм, сперматозоид должен слиться с яйцеклеткой в одну клетку. Этот процесс называют оплодотворением, а образовавшуюся клетку — зиготой. После оплодотворения зигота делится на две клетки. Потом из этих двух клеток образуется четыре, из четырех — восемь, из восьми — шестнадцать и так далее. Количество клеток стремительно уве-личивается, из них формируется зародыш (рис. 35).
У одних животных (например, млекопитающих — коров, со-бак, коз) зародыш находится в теле матери. Питательные веще-ства и кислород он получает из ее организма. У других (насеко-мые, большинство рыб, змей, птицы) зародыш развивается не в организме матери, а в яйце. Яйца рыб и лягушек называют ик-рой. Яйцо содержит запас питательных веществ, который заро-дыш использует для развития и роста.
Через определенное время животное рождается , то есть ос-тавляет материнское тело или яйцо. В наследство ему достались признаки и свойства родителей. Вот почему размножение назы-вают процессом воспроизведения себе подобных.
В живой природе рассмотренный вид размножения является самым распространенным и называется половым размножением , поскольку в нем принимают участие половые клетки — яйцеклет-ка и сперматозоид.
Некоторые животные могут также размножаться частями тела . Например, из отломанного луча морской звезды (рис. 36) или из отдельного участка тела дождевого червя вырастает новый ор-ганизм.
Для того чтобы мужская половая клетка слилась с женской, пыльца должна попасть на рыльце пестика. Перенесение пыль-цы на рыльце пестика называют опылением .
Многим растениям свойственно так называемое перекрест-ное опыление . Оно состоит в том, что пыльца с тычинки цветка одного растения попадает на рыльце пестика другого цветка такого же растения. Хорошо вам известным клеверу, гречихе, белой акации, яблоне в этом помогают насекомые. Отсюда и название — насекомые-опылители.
Цветки растений, которым свойственно перекрестное опы-ление, имеют преимущественно яркую окраску и душистый аромат. Как вы считаете, какое это имеет значение дли их опыления?
У ржи, дуба, березы, ольхи и некоторых других растений цветки мелкие, непоказные и без запаха. Перекрестное опыле-ние этих растений осуществляется при помощи ветра. Сухую и легкую пыльцу он разносит на большие расстояния.
Попав на рыльце пестика, пыльца прорастает внутрь через столбик пестика, доставляя мужские половые клетки к яйце-клетке. Осуществляется оплодотворение. Из цветка развива-ется плод с одним (персик, вишня и т. п.) или многими (огу-рец, яблоко) семенами. Плоды бывают сухими и сочными, имеют различную форму (рис. 38). Их предназначение — защи-щать семя.
В чем заключается особенность семени? В том, что оно содер-жит зародыш и достаточный запас питательных веществ, для того чтобы при благоприятных условиях зародыш пророс — превратился в молодое растение.
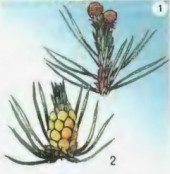 |
| Рис. 39. Шишки сосны: 1 - женская; 2 - мужская |
Встречается у растений и самоопыление, как, например, у проса, фасоли, гороха. При таком опылении пыльца попадает на рыльце пестика своего же цветка и происходит оплодотворение. У сосны, ели, можжевельника цветков нет. Но имеются муж-ские и женские шишки. В них созревают половые клетки. Их опылению также способствует ветер (рис. 39).
Кроме полового у растений бы-вает и бесполое размножение. Его название уже указывает на то, что оно происходит без участия половых клеток. Это размножение осу-ществляется с помощью других клеток тела, в том числе частями растения (рис. 40).
Размножение растений частями их тела называют вегетативным. Оно достаточно распространено. Например, картофель размножают клубнями, клубнику — усами, ма-лину — корневыми побегами, тюль-пан — луковицами, ландыш — корневищами. Достаточно посадить один куст клубники, и через год количество ее кустов достигнет почти сотни. Материал с сайта
Вегетативное размножение частями стебля — черенками, напри-мер смородины или винограда, часто применяется садоводами. Во влажной почве черенки образуют корни и дают начало новому растению. В природе вегетативное размножение способствует быстро-му расселению растений на значи-тельные территории.
Существуют другие способы раз-множения организмов. Так, грибам свойственно бесполое размножение спорами . Вспомните, стоит проко-лоть зрелый гриб-дождевик, как над ним появится коричневое облачко.
Это и есть его споры. Например, во-доросли, мхи, хвощи, папоротники размножаются не только половым путем, но и с помощью спор.
Одноклеточные организмы не имеют специальных половых органов. Поэтому они размножаются преимущественно делением клетки пополам (рис. 41).
На этой странице материал по темам:
Как размножаются животнообразные одноклеточные организмы
Реферат на тему гетативное размножение корнем и рисунок
Как размножаются животные 5 класс доклад
Доклад как размножаются растения 2 класс
Названме организмп, половой/бесполый
Вопросы по этому материалу:
Размножение, присущее всем организмам свойство воспроизведения себеподобных, обеспечивающее непрерывность и преемственность жизни. В основе всехформ размножения у организмов, обладающих клеточным строением, лежит делениеклетки. Предлагались различные классификации форм размножения. Основныхспособов размножения три: бесполое, вегетативное и половое. При бесполом размножениеорганизм развивается из одной клетки, не дифференцированной в половомотношении. При вегетативном размножение начало новому организму даютмногоклеточные зачатки, иногда сложно дифференцированные. Половому размножениюпредшествует образование гамет (половых клеток); само размножение сводится к ихслиянию в зиготу - оплодотворению, сопровождающемуся объединением не толькоцитоплазмы гамет. Начало периода в одних случаях совпадает с прекращениемроста, в других - не влечёт за собой остановки роста индивидуума и прекращаетсятолько с наступлением старости или продолжается до смерти организма, в третьих- начинается через несколько лет после прекращения роста. Размножение бываетоднократным или многократным. Для одноклеточных организмов, размножающихсяделением, а также для однолетних и двулетних цветковых растений размножение одновременноявляется завершением их жизненного цикла. Некоторые (так называемыемонокарпические) многолетние растения, а также немногие виды рыб размножаются 1раз в жизни.
Значительно чаще в растительном и животном мире наблюдаетсямногократное размножение. Каждому виду свойственна определённая интенсивностьразмножения, меняющаяся иногда в довольно широких пределах в зависимости отусловий существования.
/>Размножение животных. Бесполое размножение простейших происходит путём деления надвое (поперечно илипродольно). У некоторых из них продукты деления не разъединяются и в результатевозникают колонии. Кроме деления надвое, существуют и другие формы бесполого размноженияпростейших: множественное деление, или шизогония, и ряд других
Вегетативное размножение многоклеточных возникло вторично инезависимо в разных группах организмов и осуществляется в самых различныхформах. Его часто объединяют с размножением при помощи одноклеточных зачатковпод названием бесполого размножения (в широком смысле слова) по признакуотсутствия полового процесса, хотя по происхождению это две различные формыразмножений. Среди многоклеточных животных способностью к вегетативному размножениюобладают преимущественно низшие - губки, кишечнополостные, плоские черви,мшанки, некоторые кольчецы. Среди хордовых вегетативное размножение распространеноу вторично упрощённых форм - оболочников. Оно осуществляется чаще почкованием(наружным или внутренним), реже - делением тела на равные участки. Укишечнополостных и мшанок незавершённое вегетативное размножение приводит кобразованию колоний.
При половом размножение основной процесс - слияние гамет. Приэтом в зиготе объединяется несущий наследственную информацию хромосомныйкомплекс, происходящий от обоих родителей. Возникновение полового процесса наоснове более примитивного бесполого размножение явилось в эволюциипрогрессивным фактором, повысившим наследственную изменчивость и,соответственно, темп эволюции. Гаметы всегда гаплоидны - несут одинарный наборхромосом. Зигота диплоидна - обладает парным набором хромосом. Преобразованиедиплоидного хромосомного комплекса в гаплоидный осуществляется в результате мейоза.Последний у многоклеточных животных предшествует образованию гамет. Упростейших место его по ходу жизненного цикла может быть различным. У некоторыхпростейших имеет место изогамия - копуляция морфологически неразличимых гамет.У других наблюдается более или менее резко выраженная анизогамия - наличиеразличных гамет, из которых одни - женские, или макрогаметы, крупны и богатыцитоплазмой и резервными веществами, тогда как другие - мужские, илимикрогаметы, очень мелки и подвижны. Крайняя форма анизогамии - оогамия, прикоторой макрогамета представлена крупной, неподвижной, богатой резервнымивеществами яйцевой клеткой, а микрогаметы - подвижными мелкими сперматозоидами.
У некоторых животных (многие членистоногие, особеннонасекомые) развитие половой клетки в определённых условиях происходит безоплодотворения. Эта вторично упрощённая форма полового размножения называется партеногенезом,или девственным размножением. Особую его форму представляет педогенез -девственное размножение на личиночной стадии (свойственное некоторым двукрылыми жукам).
Для многих животных характерно закономерное чередованиеразных форм размножений, которое может сочетаться с чередованием морфологическиразличных поколений. Различают первичное и вторичное чередование поколений. Припервичном чередуются бесполое и половое размножение. Это наблюдается у многихпростейших (например, у споровиков). К вторичной форме чередования поколенийотносятся метагенез и гетерогония. При метагенезе чередуются половое размножениеи вегетативное размножение; так, в классе гидроидных (тип кишечнополостных)полипы почкуются и образуют колонии, на которых развиваются медузы (половоепоколение); последние отделяются от колоний, свободно плавают в воде, у нихразвиваются половые железы. Пример гетерогонии - чередование поколений уветвистоусых ракообразных и коловраток. Большую часть лета эти животныеразмножаются партеногенетически, лишь к осени у них развиваются самцы и самки.
На наступление периода размножения и его интенсивность большоевлияние оказывают условия среды - температура, длина светового дня,интенсивность освещения, питание и т.п. У высших животных деятельностьорганов размножения связана с функциями эндокринных желёз, что позволяетстимулировать или задерживать половое созревание. Например, у рыбдополнительная пересадка гипофиза или введение его гормонов вызываетнаступление половозрелости, что используется в практике разведения ценных рыб,например осетровых.
/>Размножение растений. Для растений наряду споловым, характерно многообразие способов бесполого и вегетативное размножение.Вегетативное размножение осуществляется путём развития новых особей извегетативных органов или их частей, иногда из особых образований, возникающихна стеблях, корнях или листьях и специально предназначенных для вегетативного размножения. Как у низших растений, так и у высших способы вегетативного размножения разнообразны.У высших растений в его основе лежит способность к регенерации. Вегетативное размножениеиграет очень большую роль в природе и широко используется человеком. Многиекультурные растения размножают почти исключительно вегетативным путём - лишь вэтом случае сохраняются их ценные сортовые качества.
Бесполое размножение многих растений осуществляется припомощи образования подвижных или неподвижных спор. У низших растений образуютсяспециальные споры бесполого размножения, которые возникают эндогенно - обычновнутри особых спорангиев (у водорослей и низших грибов) или экзогенно - наповерхности ответвлений таллома - конидиеносцев (у высших грибов). У растений,связанных в своём развитии с водной средой, эти споры подвижные. Спорообразованиеу высших растений (кроме семенных) - обязательная фаза их жизненного цикла,правильно чередующаяся с половым размножением. Половое размножение имеется убольшинства растений; отсутствует оно у синезелёных водорослей, многиенесовершенных грибов, лишайников. У синезелёных водорослей полового размножения,по-видимому, никогда не было, у несовершенных грибов и лишайников оно,вероятно, утрачено в процессе эволюции. У остальных низших растений половое размножениевыражено крайне разнообразно. В результате полового процесса (конъюгация,изогамия, гетерогамия, оогамия, гаметангиогамия) у них образуется зигота,которая переходит в состояние покоя (у большинства зелёных водорослей,некоторых бурых водорослей и у низших грибов) или немедленно прорастает, даётлибо диплоидный вегетативный таллом (у большинства бурых водорослей), либоспоры полового размножения (карпоспоры красных водорослей). У сумчатых ибазидиальных грибов половой процесс своеобразен: типичная зигота у них необразуется, начальный этап размножения (слияние протоплазмы) отделен некоторымпромежутком времени от конечного (слияние ядер), за которым следует образованиеаскоспор или базидиоспор. Для грибов характерно образование двуядерногомицелия, который у базидиальных грибов составляет основу и вегетативного тела(грибницы) и плодовых тел. Низшие растения, образующие много спор бесполого размножения,обычно обладают невысокой энергией полового размножения. У мхов органы половогоразмножения возникают на самом растении - гаметофите (половое поколение). Уодних мхов мужские половые органы (антеридии) и женские (архегонии) развиваютсяна одном и том же растении, у других - на разных. В архегонии находится однакрупная яйцеклетка. В антеридии развивается множество подвижных сперматозоидов.В каплях росы или дождя сперматозоиды, вышедшие из антеридия, достигаютархегония, проникают внутрь его и сливаются с яйцеклеткой. Из оплодотворённойяйцеклетки развивается спорогоний, внутри которого путём мейоза развиваютсяспоры для бесполого размножения. У папоротников, хвощей, плаунов, селагинеллорганы полового размножения сходны с таковыми мхов, но упрощены и образуются намаленьком заростке (гаметофите), развивающемся из споры и живущем у большинстваиз них независимо от спорофита. Заростки обычно однополые, у некоторых видов -обоеполые. Оплодотворение такое же, как у мхов.
Семенным растениям свойствен особый тип размножения -семенное, при котором формируются семена - зачатки, обеспечивающие наиболееэффективное расселение вида. У голосеменных семена развиваются из семяпочек,большей частью на особых видоизменённых листьях - спорофиллах (споролистиках).В семяпочке, которая гомологична мегаспорангию, возникают 4 мегаспоры, 3 из нихотмирают, а оставшаяся путём деления даёт заросток, состоящий из комплексатонкостенных клеток - эндосперма и 2 или нескольких примитивных архегониев. Изоплодотворённых яйцеклеток архегониев развиваются зародыши, а из семяпочки -семя, содержащее 1 зародыш (остальные отмирают). У покрытосеменных растенийсемена развиваются из семяпочек, заключённых внутри завязи цветка. Внутрисемяпочки также образуются мегаспоры. У большинства растений 3 из них обычноотмирают, а оставшаяся даёт зародышевый мешок, состоящий обычно из 7 клеток,одна из которых - яйцеклетка - после оплодотворения развивается в зародыш. Изсемяпочки образуется семя, а вся завязь превращается в плод. У некоторыхцветковых растений семена образуются без оплодотворения.
Половое размножение , различные видыразмножения животных и растительных организмов, при которых новый организмразвивается обычно из зиготы, образующейся в результате оплодотворения, т. е.слияния женских и мужских половых клеток - гамет. К половому размножениюотносят и партеногенетическое размножение, при котором новый организмразвивается из неоплодотворённой женской половой клетки. Половое размножениехарактерно для представителей всех типов животных и растений, оно неустановлено достоверно только у бактерий и синезелёных водорослей. Взависимости от формы, относительной величины и подвижности сливающихся гаметразличают следующие виды полового размножения, или полового процесса: изогамию,анизогамию, гетерогамию, оогамию.
Оплодотворение , сингамия, у растений,животных и человека - слияние мужской и женской половых клеток - гамет, врезультате чего образуется зигота, способная развиваться в новый организм.Оплодотворение лежит в основе полового размножения и обеспечивает передачунаследственных признаков от родителей потомкам.
/>Оплодотворение у растений .Оплодотворение свойственно большинству растений; ему обычно предшествуетобразование гаметангиев - половых органов, в которых развиваются гаметы. Частоэти процессы объединяют под общим названием половой процесс. Растения, имеющиеполовой процесс, имеют в цикле развития и мейоз, т. е. обнаруживают сменуядерных фаз. Типичного полового процесса нет у бактерий и синезелёныхводорослей; неизвестен он и у некоторых грибов. Типы полового процесса у низшихрастений разнообразны. У ряда зелёных водорослей он может осуществляться безобразования гамет, в результате слияния двух одноклеточных организмов. Слияниеимеющих жгутики гамет, форма и размеры которых одинаковы, называется изогамией />(см.рис. 1 , 1). Этот тип полового процесса присущмногим водорослям. Одноклеточные водоросли (например, некоторые хламидомонады)как бы сами превращаются в гаметангии, образуя гаметы; у многоклеточныхгаметангиями становятся некоторые клетки, не отличающиеся от других (например,у улотрикса, ульвы), или возникают морфологически отличные гаметангии(например, у эктокарпуса). Многие изогамные водоросли гетероталличны: сливаютсялишь физиологически различные (+ и –) гаметы. Для водорослей конъюгат(например, спирогиры) характерна конъюгация: протопласт одной клетки перетекаетв другую (принадлежащую той же или дРазмножение особи), сливаясь с еёпротопластом />(рис. 1 , 2). Слияние имеющих жгутикигамет различной величины (большая - женская, меньшая - мужская; например, унекоторых хламидомонад) называется гетерогамией />(рис. 1 ,3). Слияние крупной безжгутиковой женские гаметы (яйцеклетка) и мелкой мужской,чаще имеющей жгутики (сперматозоид), реже - безжгутиковой (спермаций),называется оогамией. Женские гаметангии большинства оогамных низших растенийназываются оогониями, мужские - антеридиями. Оогамия характерна для многихзелёных, диатомовых, бурых />(рис. 1 , 4) и всехкрасных водорослей, некоторых низших грибов. У гологамных, изо-, гетеро- имногих оогамных растений оплодотворение происходит в воде, у некоторыхоогамных (вольвокса, вошерии) - в женских гаметангиях - оогониях, к которымвышедшие в воду сперматозоиды активно перемещаются (что, видимо, обусловлено хемотаксисом),а спермации красных водорослей - пассивно, током воды. У растений сгаметангиогамией гаметы не дифференцируются. Так, у мукоровых грибов сливаютсямногоядерные гаметангии, возникающие на концах выростов мицелиев (разных пригетероталлизме) />(рис. 1 , 5); при этом попарносливаются и ядра. Этот тип гаметангиогамии называется зигогамией. У большинствасумчатых грибов многоядерный протопласт антеридия переливается в базальнуюклетку женского гаметангия (аскогон), содержащую протопласт со множеством ядер;ядра лишь попарно сближаются, образуя т. н. дикарионы (первый этап половогопроцесса - плазмогамия). Из аскогона вырастают гифы, в них ядра дикарионовсинхронно делятся; на концах гиф возникают сумки - клетки, содержащие подикариону. В сумках (асках) происходит второй этап полового процесса - кариогамия,т. е. слияние ядер />(рис. 1 , 6). Для базидиальныхгрибов характерна соматогамия: они не образуют ни гамет, ни гаметангиев;плазмогамия происходит у них при слиянии двух одноядерных клеток, т. н.первичных (+ и –) мицелиев; возникающая при этом двуядерная клетка даёт началовторичному мицелию, состоящему из клеток, содержащих дикарионы; на этом мицелииобразуются базидии, в них и происходит кариогамия />(рис.1 , 7). Гаметангио- и соматогамия - выработанное грибами в процессе эволюцииприспособление к существованию вне водной среды.
Все высшие растения оогамны, но оплодотворение у нихосуществляется по-разному. Типичные гаметангии высших растений - антеридии(мужские) и архегонии (женские) многоклеточны; клетки наружного слоя гаметангиястерильны. Яйцеклетки образуются в архегониях по одной, сперматозоиды - вантеридиях, как правило, помногу. Мохо- и папоротникообразным для осуществленияоплодотворения необходима вода, в которой вышедшие из антеридиев сперматозоидыплывут к архегониям. Из вскрывшейся вершины готового к оплодотворению архегониявыступает слизь, привлекающая сперматозоиды. Двигаясь в слизи, сперматозоидыдостигают яйцеклетки и один из них сливается с ней />(рис.1 , 8 и 9). У папоротникообразных и семенных растений оплодотворениепроисходит на (или в) заростке (гаметофите), существующем у первыхсамостоятельно, а у вторых - на спорофите. У равноспоровых папоротниковзаростки обоеполы, у разноспоровых и всех семенных растений раздельнополы. Усеменных растений антеридиев нет: сперматозоиды (у саговников, гинкго) илибезжгутиковые спермии (у всех остальных) образуются в мужских заростках (пыльцевыхзёрнах). У некоторых голосеменных (гнетум, вельвичия) и всех покрытосеменныхархегониев нет и яйцеклетки находятся в женских заростках. У семенных растенийоплодотворение возможно лишь после опыления - перенесения пыльцевых зёрен измикроспорангиев в пыльцевые камеры семезачатков (у голосеменных) или на рыльцапестиков (у покрытосеменных). У саговников и гинкго сперматозоиды выходят вархегониальную камеру семезачатка />(рис. 1 , 10) и,двигаясь в жидкости, выработанной самим растением, достигают архегониев. Усеменных растений, имеющих спермии, последние перемещаются к яйцеклеткам попыльцевым трубкам />(рис. 1 , 11 и 12). Упокрытосеменных происходит двойное оплодотворение: один спермий сливается сяйцеклеткой, второй - с центральной клеткой зародышевого мешка (женского заростка).Осуществление оплодотворения вне зависимости от наличия свободной воды - одноиз важнейших приспособлений семенных растений к существованию на суше.
/>Оплодотворение у животных ичеловека заключается в слиянии (сингамии) двух гамет разного пола - спермия(сперматозоида) и яйца. Оплодотворение имеет двоякое значение: 1) контактспермия с яйцом выводит последнее из заторможенного состояния и побуждает кразвитию; 2) слияние гаплоидиых ядер спермия и яйца - кариогамия - приводит квозникновению диплоидного синкариона, объединяющего отцовские и материнскиенаследственные факторы. Возникновение при оплодотворении новых комбинаций этихфакторов создаёт генетическое разнообразие, служащее материалом дляестественного отбора и эволюции вида. Необходимая предпосылка оплодотворения -уменьшение числа хромосом вдвое, что происходит во время мейоза. Эти деления умужских гамет осуществляются до формирования спермия, тогда как соотношениямежду делениями созревания яйца и оплодотворениия у разных животных различно:спермий может проникать в яйцо до начала мейоза (губки, некоторые черви,моллюски, из млекопитающих - собака, лисица, лошадь); на стадии метафазы 1-годеления созревания (некоторые черви, моллюски, насекомые, асцидии); на стадииметафазы 2-го деления (ланцетник, многие позвоночные) и после завершения мейоза(кишечнополостные, морские ежи).
Встреча сперматозоида с яйцом обычно обеспечиваетсяплавательными движениями мужских гамет после того, как они выметаны в воду иливведены в половые пути самки. Встрече гамет способствует выработка яйцами гамонов,усиливающих движения спермиев и продлевающих период их подвижности, а такжевеществ, вызывающих скопление спермиев вблизи яйца. Возникновение такихскоплений у гидроидных полипов из рода Campanularia и некоторых рыб иногдарассматривают как следствие привлечения спермиев (хемотаксис), подобногонаблюдаемому при оплодотворении у мхов, папоротников и др. Однако существованиенаправленных движений для спермиев животных недоказано; сперматозоиды двигаютсябеспорядочно и вступают в контакт с яйцом в результате случайного столкновения,а образование их скоплений, вероятно, вызывается действием механизма типа«ловушки», задерживающего спермии, случайно приблизившиеся к яйцу.
Зрелое яйцо окружено оболочками, имеющими у некоторыхживотных отверстия для проникновения спермиев - микропиле. У большинстваживотных микропиле отсутствует, и, чтобы достигнуть поверхности ооплазмы,спермии должны проникнуть через оболочку, что осуществляется с помощьюспециального органоида сперматазоида - акросомы. После того как спермий концомголовки коснётся яйцевой оболочки, происходит акросомная реакция: акросомараскрывается, выделяя содержимое акросомной гранулы />(рис.2 , стадии I, II), и заключённые в грануле ферменты растворяют яйцевыеоболочки. В том месте где раскрылась акросома, её мембрана сливается сплазматической мембраной спермия; у основания акросомы акросомная мембранавыгибается и образует один или несколько выростов />(рис.2 , стадия II) которые заполняются расположенным между акросомой и ядром(субакросомальным) материалом, удлиняются и превращаются в акросомные нити илитрубочки />(рис. 2 , стадия III). Длина этих нитей уразных животных варьирует от 1 до 90 мкм (в зависимости от толщины барьера,который спермию приходится преодолевать). Акросомная нить проходит черезрастворённую зону яйцевой оболочки, вступает в контакт с плазматическоймембраной яйца и сливается с ней />(рис 2 , стадииIII, IV). У животных, спермии которых проникают в яйцо через микропиле(насекомые, головоногие моллюски, осетровые и костистые рыбы), акросомаутрачивает своё первоначальное значение и иногда редуцируется или полностьюисчезает (у некоторых веснянок, костистых рыб). У млекопитающих овулировавшееяйцо, кроме оболочки, окружено несколькими слоями фолликулярных клетокяйценосного бугорка. У лошади, коровы, овцы фолликулярные клетки рассеиваютсявскоре после овуляции, и спермий свободно достигает поверхности яйцевойоболочки. У большинства млекопитающих клетки яйценосного бугорка сохраняются напротяжении нескольких часов и, чтобы проникнуть через этот барьер, спермиивыделяют фермент гиалуронидазу, который растворяет вещество, связывающеефолликулярные клетки между собой. Гиалуронидаза, как и фермент, растворяющийяйцевую оболочку, заключена в акросоме. Сразу после эякуляции спермиинеспособны к выделению этих ферментов; такая способность возникает поддействием содержимого женских половых путей, вызывающего определённыефизиологические изменения спермиев (процесс капацитации).
С момента слияния плазматических мембран гамет в местеконтакта акросомной нити с поверхностью ооплазмы яйцо и спермий - уже единаяклетка - зигота. Вскоре обнаруживаются первые признаки активации яйца: кортикальнаяреакция и стягивание ооплазмы в месте контакта с акросомной нитью спермия,приводящее к образованию воспринимающего бугорка. Ооплазма этого бугоркаобтекает ядро, центриоли и митохондрии сперматозоида, а иногда и осевойстержень его хвоста, вовлекая их в глубь яйца, тогда как плазматическаямембрана спермия остаётся на поверхности и встраивается в плазматическуюмембрану яйца, так что поверхностная мембрана зиготы имеет мозаичное строение.Погрузившись в ооплазму, головка спермия поворачивается на 180°, и у её основанияформируется сперматическая звезда />(рис. 3 , стадииI, II). Постепенно головка набухает и преобразуется в пузыревидный мужскойпронуклеус, перемещающийся вслед за сперматической звездой, которая как быувлекает его за собой />(рис. 3 , стадии II, III, IV).Мужской пронуклеус сближается с женским, а сперматическая звезда делится надве, участвующие затем в образовании веретена 1-го деления дробления. Укишечнополостных, плоских червей, морских ежей пронуклеусы сливаются в единоеядро зиготы />(рис. 3 , стадия V), у некоторыхкруглых червей, моллюсков, ракообразных, рыб и земноводных они длительное времяостаются в тесном контакте, но не сливаются, и объединение отцовского иматеринского ядерного материала происходит только на стадии метафазы 1-годеления дробления. Одновременно с этими изменениями в яйце повышаетсяинтенсивность обмена веществ: увеличивается проницаемость клеточной мембраны,активируется синтез белка и дРазмножение
При оплодотворении яиц животных с наружным осеменением вяйцо проникает только один спермий (физиологическая моноспермия); этообеспечивается особым механизмом, в основе которого лежит процесс секрециисодержимого кортикальных телец, предотвращающий проникновение спермиев вооплазму. Среди животных с внутренним осеменением наряду с моноспермнымивстречаются и такие, у которых в яйцо проникает несколько спермиев(физиологическая полиспермия); однако и в этих случаях с женским пронуклеусомсливается только одно сперматическое ядро.
Рис. 1. Оплодотворение у растений: 1 - изогамия у хламидомонадыРейнхардта; 2 - конъюгация у спирогиры; 3 - гетерогамия у хламидомонады Брауна;4 - оогамия у фукуса: а - оогоний, б - группа антеридиев, в - сперматозоид, г -сперматозоиды близ яйцеклетки; 5 - зигогамия у мукора; 6 - дикарион (а),диплоидное ядро (б) и образование спор (в) в сумке сумчатого гриба; 7 -дикарион (а) и диплоидное ядро (б) в базидии и образование спор (в) убазидиального гриба; 8 - вскрывшийся архегоний (а), выход сперматозоидов изантеридия (б) и сперматозоиды (в) у зелёных мхов; 9 - выход сперматозоидов изантеридия (а), сперматозоид (б) и вскрывшийся архегоний (в) у равноспоровыхпапоротников; 10 - пыльцевое зерно (а), сперматозоид (б) и часть семезачаткасаговника (в), в котором видны часть женского заростка с архегониями и мужскиезаростки, выдающиеся в архегониальную камеру; 11 - пыльцевое зерно (а) и частьсемезачатка сосны (б), где видны часть женского заростка с архегониями ипыльцевая трубка со спермиями; 12 - пыльцевое зерно (а) и часть зародышевогомешка покрытосеменного растения (б): ядро одного спермия находится близ ядраяйцеклетки, а ядро другого - близ вторичного ядра зародышевого мешка.
Рис. 2. Схема осуществления акросомной реакции: А - у кольчатогочервя; Б - у кишечнодышащего; В - у морского ежа; I-IV - последовательныестадии реакции; а - акросомная мембрана, аг - акросомная гранула, ан -акросомная нить, с - плазматическая мембрана спермия, см - субакросомальныйматериал, я - плазматическая мембрана яйца.
Рис. 3. Оплодотворение у морского ежа: I-IV - последовательныестадии преобразования спермия и формирования мужского пронуклеуса; V - слияниепронуклеусов; з - сперматическая звезда, м - митохондрия из средней частиспермия, п - женский пронуклеус, пт - полярные тельца, х - хвостовой отделспермия, я - ядро спермия.
Половое созревание , пубертатный период (отлат. pubertas - половая зрелость), период в индивидуальном развитии животногоорганизма, в течение которого путём глубоких внутренних перестроек он достигаетполовой зрелости (способности к размножению). У человека половое созреваниехарактеризуется ускорением роста отдельных сегментов скелета с последующимустановлением окончательных пропорций тела, завершением формирования вторичныхполовых признаков, половых органов. Существенный признак полового созревания -выделение специфических продуктов внешней и внутренней секреции половых желёз,что проявляется у девушек установлением менструального цикла, у юношей - эякуляциями(вначале обычно при поллюциях). П. с. охватывает процессы перестройки гормональнойрегуляции, физического развития и психической сферы. Так, в последнейпроисходит становление полового сознания, т. е. способности человека не толькоосознавать себя носителем определённого пола, но и регламентировать своёсексуальное поведение в соответствии с системой принятых в данном обществеморально-этических установок. В формировании полового сознания условно можновыделить несколько фаз: элементарных представлений о половых различиях безспецифической чувственной окраски; платонической влюблённости; пробужденияспецифических сексуальных эмоций, направленных на генитальную сферу; фазузрелого полового сознания - гармоничного соотношения всех его компонентов.
Начало и темп полового созревания определяютсявзаимодействием конституциональных факторов и влияний внешней среды. Возрастныерамки полового созревания подвержены широким индивидуальным колебаниям и (сучётом процессов акцелерации) укладываются в период: у девочек - от 8-9 до16-17 лет, у мальчиков - от 10-11 до 19-20 лет. Неблагоприятные бытовые условияи дРазмножение влияния (неполноценное питание, перенесённые заболевания)проявляются задержкой, ускорением (реже) и дисгармониями физического,психического и полового развития.
Половая жизнь , совокупностьсоматических (телесных), психических и социальных процессов и отношений, воснове которых лежит и посредством которых удовлетворяется половое влечение.
/>Пол и половое поведение .Общей биологической основой полового поведения является инстинкт продолжениярода; его конкретные формы (разделение функций между полами, особенностирепродуктивного цикла, ритуал ухаживания, техника полового акта и т.д.)различаются в зависимости от вида и рода, а также пола, возраста и условийжизни организма. Наряду с инстинктивной, генетически заданной организму программойнормальное половое поведение обеспечивается посредством научения, через общениесо сверстниками, родителями и др. Чем выше уровень организации и психикиживотного, тем большую роль в его поведении играют усвоенные и выработанные впроцессе индивидуального развития навыки. Например, морские свинки, выращенныев условиях полной изоляции, будучи по достижении половой зрелости подсажены кособям противоположного пола, в большинстве случаев проявляют нормальнуюполовую активность, инстинктивно находя адекватные своему виду формы спариванияи т.д. В отличие от этого, макаки-резусы, выращенные в изоляции,оказываются неспособными к нормальной половой жизни, т.к. у них невыработаны необходимые психологические механизмы общения, Чем сложнее животное,тем глубже и многограннее связь его полового поведения с др. аспектамижизнедеятельности. Половое поведение животных необходимо рассматривать вединстве индивидуально-генетических и групповых (формирующихся и проявляющихсятолько в общении с др. особями того же вида) характеристик.
У человека к этим факторам добавляется и играет важнейшуюроль ряд социально-культурных детерминант. Биологический пол индивида включаетнесколько относительно автономных компонентов, последовательно формирующихся впроцессе развития организма: 1) хромосомный (генетический) пол, которыйопределяется в момент зачатия; 2) гонадный пол - формирование мужских илиженских половых желёз; 3) гормональный пол зародыша, от которого зависитдифференциация гениталий - половых органов (гормоны также сильно влияют напсихику); 4) морфологический пол - строение внутрирепродуктивных органов инаружных гениталий; 5) пубертатный, т. е. связанный с половым созреванием,гормональный пол, ответственный за появление вторичных половых признаков, и др.Нарушение любого из звеньев этой системы так или иначе сказывается на половойфункции. Однако сам по себе биологический пол ещё не превращает индивида вмужчину или женщину и не гарантирует адекватного полового поведения. Для этогонеобходима также соответствующая психосексуальная идентификация(отождествление): человек должен осознать свою половую принадлежность и усвоитьсоответствующую, мужскую или женскую, роль.
Психосексуальная идентификация - один из важнейших аспектовформирования личности; она осуществляется в процессе жизнедеятельности, подвлиянием воспитания и общения. Определив по внешним признакам полноворождённого (т. н. аскриптивный, приписанный пол), родители и др. взрослыесистематически воспитывают ребенка в соответствующем духе. Уже к 1,5-2 годамребёнок обычно знает свой пол и в дальнейшем основывает на этом определённыйстиль жизни и уровень притязаний, поскольку все культуры так или иначеразличают «мужские» и «женские» роли и качества. Степень соответствия своеготела и поведения стереотипу мужчины или женщины является своеобразнойпсихологической осью, вокруг которой структурируется образ собственного Я и откоторой сильно зависит самооценка и самоуважение. Позже, по мере половогосозревания, у подростка появляются соответствующие сексуальные интересы,влечение к противоположному полу. Огромное влияние оказывают при этом среда исверстники. В норме биологические и социально-психологические характеристикипола совпадают, идут в одном направлении. Однако бывают случаи, когда онирасходятся (ошибочное определение пола младенца в связи с недоразвитостью егогениталий или сомнения относительно половой принадлежности подростка в связи сзадержкой полового созревания и т.д.). Предсказать направлениепсихосексуальной идентификации и психосексуальных ориентаций личности в этихслучаях трудно; во всяком случае, социальные факторы здесь не менее важны, чемгенетические. Половое поведение человека - продукт совместного действиябиогенетических и социальных (аскриптивный пол и общественная системадифференциации мужских и женских ролей) сил.
/>Культурно-исторические аспектыполовой жизни . Одна из особенностей человеческой сексуальности состоит втом, что переживания, связанные с удовлетворением половой потребности, отделеныот её первоначальной биологические цели - продолжения рода. Эта автономиярекреативной функции (получение наслаждения) половой жизни от репродуктивной(детопроизводство) создала принципиальную возможность гуманизации половогочувства, превращения элементарного полового влечения в человеческую любовь, когдавзаимоотношения между мужчиной и женщиной служат не только средствомпродолжения рода и источником чувственного наслаждения, но и дают высочайшеесчастье глубокой психологической, душевной близости. Отсюда также вытекаетгромадное многообразие форм человеческой сексуальности. Не говоря уже оразличных замещающих (викарирующих) формах П. ж., когда оргазм вызывается путёмискусственного возбуждения гениталий, без полового акта, и которые присущи нетолько человеку, но и многим животным, человеческое воображение способноэротизировать, делать сексуально значимым практически любой реальный илиидеальный объект. Причём это происходит не только на уровне индивидуального, нои на уровне общественного сознания половой жизни - неотъемлемая составнаячасть культуры, которая, с одной стороны, выражает, обобщает и символизирует, ас другой - направляет и координирует многообразные фактические проявлениясексуальности.
Всякое развитое человеческое общество имеет систему нормполовой морали, которая регулирует взаимоотношения полов, соотнося их сдеятельностью общественных институтов, в рамках которых осуществляетсяпродолжение рода и социализация, воспитание потомства. Уже древнейшиецивилизации знали весьма различные формы П. ж., одни из которых нормативноодобрялись и даже возводились в ранг культурных символов (например, фаллическийкульт, оргиастические ритуалы Востока, Дионисии),. а др. осуждались изапрещались. В целом П. ж. усложнялась и индивидуализировалась по мере развитиякультуры и личности. Однако процесс этот противоречив. Соотношение духовных и телесныхаспектов половой жизни, равно как и отношение к различным формам сексуальности,неодинаково в разных обществах. Древнейшим и самым универсальным табу безусловноявляется запрет кровосмешения (инцеста), но насчёт других аспектов половойжизни (добрачные и внебрачные связи, онанизм, гомосексуализм и т.д.)разные общества и культуры придерживались разных, иногда даже противоположныхнорм. Уже античные авторы различают чувственное влечение, половую страсть кобладанию (эрос) и возвышенную духовную любовь (агапе, платоническая любовь).Но при этом рекреативная сторона сексуальности настолько резко отделяется отрепродуктивной, осуществляемой в браке, что, по выражению Ф. Энгельса, «… дляклассического поэта древности, воспевавшего любовь, старого Анакреонта…безразличен был даже пол любимого существа» (Маркс К. и Энгельс Ф., Соч., 2изд., т. 21, с. 79).
Для европейского средневековья характерно двойственноеотношение к половой жизни. Официальная христианская мораль в принципеантисексуальна: всё телесное она считает низменным и греховным, видя идеал вполном воздержании; даже в браке половая жизнь допускается только ради продолжениярода. Однако на бытовом уровне средневековая культура была весьма терпима ксексуальности, видя в ней нормальный и естественный аспект человеческого бытия,свободно обсуждавшийся в быту и светском искусстве.
Раннебуржуазное общество вносит существенные коррективы вэти установки. В противовес церковному аскетизму, гуманисты реабилитируют правочеловека на чувственное наслаждение, и эротика снова занимает место вофициальной «высокой» культуре. Но вслед за тем религиозная пуританская моральсоздаёт систему антисексуальных установок, в свете которых половая жизньпредстаёт не только греховной, но и несовместимой с самореализацией личности всфере труда и накопления богатства. В жизни по принципу «делу - время, потехе -час» для сексуальности не находится места, она снова запрещается как нечтопостыдное. Дуалистическое противопоставление «возвышенной» любви «низменному»,«животному» половому влечению пронизывает и медико-педагогическую литературу18-19 вв. Если античные авторы стремились научить человека наслаждаться, топедагоги нового времени озабочены главным образом тем, как ослабить половоевлечение, оттянуть начало половой жизни Мораль полового воздержанияформулируется при этом в тех же выражениях, что и буржуазные принципы экономии,накопления, самоконтроля. Стремление «десексуализировать» человека и культуру,подкрепленное цензурной политикой (многие классические произведения литературыи искусства в своё время считались «непристойными»), увеличивает разрыв междуморальным сознанием общества и реальным поведением людей. Половая жизнь ассоциируетсяс чувством вины, уходит в «подполье», что, в свою очередь, порождает неврозы.Именно это противоречие зафиксировал в своей клинической практике и обобщилтеоретически австрийский врач З. Фрейд. Особенно тяжёлым было положение женщин,которых викторианская философия считала в принципе асексуальными, лишённымиполовых потребностей. Эта установка способствовала распространению фригидности.Для мужчин холодность официального буржуазного брака компенсироваласьузаконенной проституцией.
Кризис буржуазной системы ценностей, а также такие факторы,как рост автономии молодёжи от родителей в городской среде, ускорение половогосозревания подростков в связи с акцелерацией, вовлечение женщин вобщественно-трудовую жизнь и рост женского равноправия, ослабление традиционнойрелигиозной морали, рождение научной сексологии и распространение светского,рационального отношения к вопросам пола, появление эффективных контрацептивов(см. Контрацепция), позволяющих при правильном применении избежать нежелаемойбеременности, - всё это вместе взятое вызвало значительные сдвиги в половойжизни в сторону либерализации половой морали и ослабления «двойного стандарта»(т. е. разных норм поведения для мужчин и для женщин). Половая жизнь теперьначинается значительно раньше (в ФРГ с 1960 по 1970 у более образованной частимолодёжи возраст начала активной половой жизни снизился в среднем на 3-4 года).Отсюда - ряд социальных и этических проблем. В буржуазном обществе этот процессвыступает прежде всего отрицательными сторонами - потребительским отношением ксексу, ростом отчуждённого, коммерческого эротизма, порнографии,дезорганизацией семьи. Гипертрофия сексуальности, провозглашение её главной иопределяющей стороной человеческой жизни, типичные для некоторых «левых»теоретиков, усматривающих в «сексуальной революции» ключ ко всеобщемуосвобождению, так же вредны и несостоятельны, как её замалчивание илибиологизация. Это - две стороны одной медали. Отмечая, что в области брака иполовых отношений назревают глубокие изменения, В. И. Ленин вместе с тем резкоосуждал мелкобуржуазно-анархическую трактовку этого процесса и подчёркивалнеобходимость рассматривать его в свете коренных интересов общества, в томчисле - в связи с развитием брачно-семейных отношений, которые и при социализмеостаются важнейшим регулятором половой жизни. Отношения между полами, при всейих интимности, суть общественные отношения. Это не может не накладывать налюдей определённых обязанностей как по отношению друг к другу, так и, вособенности, по отношению к детям. Потребительское отношение к половой жизни,оборачиваясь «свободой» от ответственности и серьёзности в любви, ведёт вконечном итоге к деиндивидуализации половых отношений и эмоциональнойнеудовлетворённости. В этой области, как и в других, самоконтроль - необходимыймомент свободного самовыражения личности.
Список используемойлитературы:
1. Большая СоветскаяЭнциклопедия
2. Справочник школьника побиологии для 6-9 классов.
Реферат: Бесполое и половое размножение
Введение
Способность размножаться, т. е. производить новое поколение особей того же вида, - одна из основных особенностей живых организмов. В процессе размножения происходит передача генетического материала от родительского поколения следующему поколению, что обеспечивает воспроизведение признаков не только данного вида, но конкретных родительских особей. Для вида смысл размножения состоит в замещении тех его представителей, которые гибнут, что обеспечивает непрерывность существования вида; кроме того, при подходящих условиях размножение позволяет увеличить общую численность вида.
Каждая новая особь, прежде чем достигнуть стадии, на которой она будет способна к размножению, должна пройти ряд стадий роста и развития. Некоторые особи погибают, не достигнув репродуктивной стадии (или половозрелости) в результате уничтожения хищниками, болезней и разного рода случайных событий; поэтому вид может сохраниться лишь при условии, что каждое поколение будет производить больше потомков, чем было родительских особей, принимавших участие в размножении. Численность популяций колеблется в зависимости от баланса между размножением и вымиранием особей. Существует ряд различных стратегий размножения, каждая из которых имеет определенные преимущества и недостатки; все они будут описаны в этом реферате.
А цель моей работы: рассмотреть некоторые виды размножения.
Бесполое и половое размножение
Существуют два основных типа размножения - бесполое и половое. Бесполое размножение происходит без образования гамет, и в нем участвует лишь один организм. При бесполом размножении обычно образуются идентичные потомки, а единственным источником генетической изменчивости служат случайные мутации.
Генетическая изменчивость выгодна виду, так как она поставляет «сырье» для естественного отбора, а значит, и для эволюции. Потомки, оказавшиеся наиболее приспособленными к среде, будут обладать преимуществом в конкуренции с другими представителями того же вида и будут иметь больше шансов выжить и передать свои гены следующему поколению. Благодаря этому виды способны изменяться, т. е. возможен процесс видообразования. Повышение изменчивости может быть достигнуто путем смешения генов двух разных особей - процесса, называемого генетической рекомбинацией и составляющего важную особенность полового размножения; в примитивной форме генетическая рекомбинация встречается уже у некоторых бактерий.
Бесполое размножение
При бесполом размножении потомки происходят от одного организма, без слияния гамет. Мейоз в процессе бесполого размножения не участвует (если не говорить о растительных организмах с чередованием поколений), и потомки идентичны родительской особи. Идентичное потомство, происходящее от одной родительской особи, называют клоном. Члены одного клона могут быть генетически различными только в случае возникновения случайной мутации. Высшие животные не способны к бесполому размножению, однако в последнее время было сделано несколько успешных попыток клонировать некоторые виды искусственным образом; мы их рассмотрим в дальнейшем.
Деление
Образование спор (споруляция)
Спора-это одноклеточная репродуктивная единица обычно микроскопических размеров, состоящая из небольшого количества цитоплазмы и ядра. Образование спор наблюдается у бактерий, простейших, у представителей всех групп зеленых растений и всех групп грибов. Споры могут быть различными по своему типу и функции и часто образуются в специальных структурах.
Нередко споры образуются в больших количествах и имеют ничтожный вес, что облегчает их распространение ветром, а также животными, главным образом насекомыми. Вследствие малых размеров спора обычно содержит лишь минимальные запасы питательных веществ; из-за того что многие споры не попадают в подходящее место для прорастания, потери спор очень велики. Главное достоинство таких спор-возможность быстрого размножения и расселения видов, в особенности грибов.
Споры бактерий служат, строго говоря, не для размножения, а для того, чтобы выжить при неблагоприятных условиях, поскольку каждая бактерия образует только одну спору. Бактериальные споры относятся к числу наиболее устойчивых: так, например, они нередко выдерживают обработку сильными дезинфицирующими веществами и кипячение в воде.
Почкование
Почкованием называют одну из форм бесполого размножения, при которой новая особь образуется в виде выроста (почки) на теле родительской особи, а затем отделяется от нее, превращаясь в самостоятельный организм, совершенно идентичный родительскому. Почкование встречается в разных группах организмов, особенно у кишечнополостных, например у гидры (рис. 1), и у одноклеточных грибов, таких как дрожжи. В последнем случае почкование отличается от деления (которое тоже наблюдается у дрожжей) тем, что две образующиеся части имеют разные размеры.
Необычная форма почкования описана у суккулентного растения бриофиллум - ксерофита, часто выращиваемого в качестве декоративного комнатного растения: по краям его листьев развиваются миниатюрные растеньица, снабженные маленькими корешками (рис. 2); эти «почки» в конце концов отпадают и начинают существовать как самостоятельные растения.
Размножение фрагментами (фрагментация)
Фрагментацией называют разделение особи на две или несколько частей, каждая из которых растет и образует новую особь. Фрагментация происходит, например, у нитчатых водорослей, таких как спирогира. Нить спирогиры может разорваться на две части в любом месте.
Фрагментация наблюдается также у некоторых низших животных, которые в отличие от более высокоорганизованных форм сохраняют значительную способность к регенерации из относительно слабо дифференцированных клеток. Например, тело немертин (группа примитивных червей, главным образом морских) особенно легко разрывается на много частей, каждая из которых может дать в результате регенерации новую особь. В этом случае регенерация-процесс нормальный и регулируемый; однако у некоторых животных (например, у морских звезд) восстановление из отдельных частей происходит только после случайной фрагментации. Животные, способные к регенерации, служат объектами для экспериментального изучения этого процесса; часто при этом используют свободноживущего червя планарию. Такие эксперименты помогают понять процесс дифференцировки.
Вегетативное размножение
Вегетативное размножение представляет собой одну из форм бесполого размножения, при которой от растения отделяется относительно большая, обычно дифференцированная, часть и развивается в самостоятельное растение. По существу вегетативное размножение сходно с почкованием. Нередко растения образуют структуры, специально предназначенные для этой цели: луковицы, клубнелуковицы, корневища, столоны и клубни. Некоторые из этих структур служат также для запасания питательных веществ, что позволяет растению пережить периоды неблагоприятных условий, таких как холода или засуха. Запасающие органы позволяют растению переживать зиму и давать в следующем году цветки и плоды (двулетние растения) или выживать в течение ряда лет (многолетние растения). К таким органам, называемым зимующими, относятся луковицы, клубнелуковицы, корневища и клубни.
Зимующими органами могут быть также стебли, корни или целые побеги (почки), однако во всех случаях содержащиеся в них питательные вещества создаются главным образом в процессе фотосинтеза, происходящего в листьях текущего года. Образовавшиеся питательные вещества переносятся в запасающий орган, а затем обычно превращаются в какой-либо нерастворимый резервный материал, например крахмал. При наступлении неблагоприятных условий надземные части растения отмирают, а подземный зимующий орган переходит в состояние покоя. В начале следующего вегетационного периода запасы питательных веществ мобилизуются с помощью ферментов: почки пробуждаются, и в них начинаются процессы активного роста и развития за счет запасенных питательных веществ. Если прорастает более одной почки, то можно считать, что осуществилось размножение.
В ряде случаев образуются специальные органы, служащие для вегетативного размножения. Таковы видоизмененные части стебля - клубни картофеля, луковицы лука, чеснока, луковички в лиственных пазухах мятлика, откидыши молодила и др. Земляника размножается «усами» (рис. 3). В узлах побегов формируются придаточные корни, а из пазушных почек - побеги с листьями. В дальнейшем междоузлия отмирают, а новое растение утрачивает связь с материнским.
В практике сельского хозяйства вегетативное размножение растений используется довольно широко.
Клонирование высших растений и животных
Как уже говорилось, получение идентичных потомков при помощи бесполого размножения называют клонированием. В начале шестидесятых годов были разработаны методы, позволяющие успешно клонировать некоторые высшие растения и животных. Эти методы возникли в результате попыток доказать, что ядра зрелых клеток, закончивших свое развитие, содержат всю информацию, необходимую для кодирования всех признаков организма, и что специализация клеток обусловлена включением и выключением определенных генов, а не утратой некоторых из них. Первый успех был достигнут проф. Стюардом из Корнельского университета, который показал, что, выращивая отдельные клетки корня моркови (ее съедобной части) в среде, содержащей нужные питательные вещества и гормоны, можно индуцировать процессы клеточного деления, приводящие к образованию новых растений моркови.
Вскоре после этого Гёрдон, работавший в Оксфордском университете, впервые сумел добиться клонирования позвоночного животного. Позвоночные в естественных условиях клонов не образуют; однако, пересаживая ядро, взятое из клетки кишечника лягушки, в яйцеклетку, собственное ядро которой предварительно было разрушено путем облучения ультрафиолетом, Гёрдону удалось вырастить головастика, а затем и лягушку, идентичную той особи, от которой было взято ядро.
Такого рода эксперименты не только доказывают, что дифференцированные (специализированные) клетки содержат всю информацию, необходимую для развития целого организма, но и позволяют рассчитывать, что подобные методы можно будет использовать для клонирования позвоночных, стоящих на более высоких ступенях развития, в том числе и человека. Клонирование нужных животных, например племенных быков, скаковых лошадей и т. п., может оказаться столь же выгодным, как и клонирование растений, которое, как было сказано, уже производится. Однако применение методов клонирования к человеку сопряжено с серьезными проблемами нравственного порядка. Теоретически можно создать любое число генетически тождественных копий данного мужчины или данной женщины. На первый взгляд может показаться, что таким образом можно было бы воспроизводить талантливых ученых или деятелей искусства. Однако надо помнить, что степень влияния, оказываемого на развитие средой, еще не вполне ясна, а между тем любая клонируемая клетка должна снова пройти через все стадии развития, т. е. в случае человека-стадии зародыша, плода, младенца и т. д.
Половое размножение.
При половом размножении потомство получается в результате слияния генетического материала гаплоидных ядер. Обычно эти ядра содержатся в специализированных половых клетках - гаметах; при оплодотворении гаметы сливаются, образуя диплоидную зиготу, из которой в процессе развития получается зрелый организм. Гаметы гаплоидны - они содержат один набор хромосом, полученный в результате мейоза; они служат связующим звеном между данным поколением и следующим (при половом размножении цветковых растений сливаются не клетки, а ядра, но обычно эти ядра тоже называют гаметами.)
Мейоз - важный этап жизненных циклов, включающих половое размножение, так как он ведет к уменьшению количества генетического материала вдвое. Благодаря этому в ряду поколений, размножающихся половым путем, это количество остается постоянным, хотя при оплодотворении оно каждый раз удваивается. Во время мейоза в результате случайного расхождения хромосом (независимое распределение) и обмена генетическим материалом между гомологичными хромосомами (кроссинговер) возникают новые комбинации генов, попавших в одну гамету, и такая перетасовка повышает генетическое разнообразие. Слияние содержащихся в гаметах гаплоидных ядер называют оплодотворением или сингамией; оно приводит к образованию диплоидной зиготы, т. е. клетки, содержащей по одному хромосомному набору от каждого из родителей. Это объединение в зиготе двух наборов хромосом (генетическая рекомбинация) представляет собой генетическую основу внутривидовой изменчивости. Зигота растет и развивается в зрелый организм следующего поколения. Таким образом, при половом размножении в жизненном цикле происходит чередование диплоидной и гаплоидной фаз, причем у разных организмов эти фазы принимают различные формы.
Гаметы обычно бывают двух типов - мужские и женские, но некоторые примитивные организмы производят гаметы только одного типа. У организмов, образующих гаметы двух типов, их могут производить соответственно мужские и женские родительские особи, а может быть и так, что у одной и той же особи имеются и мужские, и женские половые органы. Виды, у которых существуют отдельные мужские и женские особи, называются раздельнополыми; таковы большинство животных и человек. Среди цветковых растений тоже есть раздельнополые виды; если у однодомных видов мужские и женские цветки образуются на одном и том же растении, как, например, у огурца и лещины, то у двудомных одни растения несут только мужские, а другие - только женские цветки, как у остролиста или у тиса.
Гермафродитизм
Партеногенез
Партеногенез - одна из модификаций полового размножения, при которой женская гамета развивается в новую особь без оплодотворения мужской гаметой. Партеногенетическое размножение встречается как в царстве животных, так и в царстве растений, и преимущество его состоит в том, что в некоторых случаях оно повышает скорость размножения.
Существует два вида партеногенеза - гаплоидный и диплоидный, в зависимости от числа хромосом в женской гамете. У многих насекомых, в том числе у муравьев, пчел и ос, в результате гаплоидного партеногенеза в пределах данного сообщества возникают различные касты организмов. У этих видов происходит мейоз и образуются гаплоидные гаметы. Некоторые яйцеклетки оплодотворяются, и из них развиваются диплоидные самки, тогда как из неоплодотворенных яйцеклеток развиваются фертильные гаплоидные самцы. Например, у медоносной пчелы матка откладывает оплодотворенные яйца (2n = 32), которые, развиваясь, дают самок (маток или рабочих особей), и неоплодотворенные яйца (n = 16), которые дают самцов (трутней), производящих спермии путем митоза, а не мейоза. Развитие особей этих трех типов у медоносной пчелы схематически представлено на рис. 4. Такой механизм размножения у общественных насекомых имеет адаптивное значение, так как позволяет регулировать численность потомков каждого типа.
У тлей происходит диплоидный партеногенез, при котором ооциты самки претерпевают особую форму мейоза без расхождения хромосом - все хромосомы переходят в яйцеклетку, а полярные тельца не получают ни одной хромосомы. Яйцеклетки развиваются в материнском организме, так что молодые самки рождаются вполне сформировавшимися, а не вылупляются из яиц. Такой процесс называется живорождением. Он может продолжаться в течение нескольких поколений, особенно летом, до тех пор пока в одной из клеток не произойдет почти полное нерасхождение, в результате чего получается клетка, содержащая все пары аутосом и одну Х-хромосому. Из этой клетки партеногенетически развивается самец. Эти осенние самцы и партеногенетические самки производят в результате мейоза гаплоидные гаметы, участвующие в половом размножении. Оплодотворенные самки откладывают диплоидные яйца, которые перезимо-вывают, а весной из них вылупляются самки, размножающиеся партеногенетически и рождающие живых потомков. Несколько партеногенетических поколений сменяются поколением, возникающим в результате нормального полового размножения, что вносит в популяцию генетическое разнообразие в результате рекомбинации. Главное преимущество, которое дает тлям партеногенез, - это быстрый рост численности популяции, так как при этом все ее половозрелые члены способны к откладке яиц. Это особенно важно в периоды, когда условия среды благоприятны для существования большой популяции, т. е. в летние месяцы.
Партеногенез широко распространен у растений, где он принимает различные формы. Одна из них - апомиксис - представляет собой партеногенез, имитирующий половое размножение. Апомиксис наблюдается у некоторых цветковых растений, у которых диплоидная клетка семязачатка-либо клетка нуцеллуса, либо мегаспора - развивается в функциональный зародыш без участия мужской гаметы. Из остального семязачатка образуется семя, а из завязи развивается плод. В других случаях требуется присутствие пыльцевого зерна, которое стимулирует партеногенез, хотя и не прорастает; пыльцевое зерно индуцирует гормональные изменения, необходимые для развития зародыша, и на практике такие случаи трудно отличить от настоящего полового размножения
Своеобразно происходит оплодотворение у цветковых растений. После оплодотворения у них из семязачатка образуется семя, содержащее зародыш и запас питательных веществ. Как же образуется в семени запас питательных веществ?
У цветковых растений происходит двойное оплодотворение. При опылении пыльцевое зерно попадает на рыльце пестика и прорастает (рис. 57 ), образуя пыльцевую трубку. Она формируется из вегетативной клетки и быстро растет, достигая завязи. В конце пыльцевой трубки находятся два спермия.
* В отличие от подвижных сперматозоидов низших растений спермии у цветковых растений неподвижны и могут проникнуть к яйцеклетке только за счет пыльцевой трубки.
Пыльцевая трубка прорастает в семязачаток, кончик ее разрывается, и спермии попадают в зародышевой мешок. Один из них сливается с яйцеклеткой. Образуется диплоидная клетка - зигота. Второй спермии сливается с диплоидным вторичным ядром зародышевого мешка. В результате образуется клетка с тройным набором хромосом, из которой путем многократных митозов формируется эндосперм - ткань, содержащая запас питательных веществ.
Двойное оплодотворение у цветковых растений


