Гаметогенез и двойное оплодотворение Цветковых растений
Двойное оплодотворение , половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon ) и рябчике (Fritillaria orientalis ). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки - эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие двойного оплодотворения - ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Оплодотворению у покрытосеменных предшествует микро- и мегаспорогенез , а также опыление .
Микроспорогенез протекает в пыльниках тычинок. При этом диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит – пыльцевое зерно .
Пыльцевое зерно снаружи покрыто двумя оболочками: экзиной и интиной . Экзина – верхняя оболочка более толстая и пропитана спороленнином – жироподобным веществом. Это позволяет пыльце выдерживать существенные температурные и химические воздействия. В экзине находятся проростковые поры, до опыления закрытые «пробочками». Интина содержит целлюлозу и эластична. В пыльцевом зерне имеются две клетки: вегетативная и генеративная .
Мегаспорогенез осуществляется в семязачатке . Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в результате остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса к интегументам, формируя зародышевый мешок . Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам: халазальному и микропилярному , и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На микропилярном полюсе находятся две синергиды и одна более крупная клетка – яйцеклетка . На халазальном полюсе располагаются антиподы . Таким образом, зрелый зародышевыый мешок содержит 7 клеток.
Опыление заключается в переносе пыльцы с тычинок на рыльце пестика.
Оплодотворение . Пыльцевые зерна, тем или иным образом попавшие на рыльце, - прорастают. Прорастание пыльцы начинается с разбухания зерна и образования пыльцевой трубки из вегетативной клетки. Пыльцевая трубка прорывает оболочку в более тонком ее месте – так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика. По мере роста пыльцевой трубки в нее переходит ядро вегетативной клетки и генеративной, которая делится и образует два спермия. Через микропиле семязачатка пыльцевая трубка проникает в зародышевый мешок, там она разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, образуя зиготу, которая затем дает начало зародышу семени. Второй спермий сливается с центральным ядром, что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Таким образом, эндосперм у покрытосеменных триплоидный и вторичный , т.к. образуется после оплодотворения.
Весь этот процесс получил название двойного оплодотворения . Он был впервые описан русским ученым С.Г.Навашиным. (1898 г.).
Антиподы и синергиды растворяются после оплодотворения, а интегументы преобразуются в семенную кожуру.
Апомиксис – развитие зародыша из неоплодотворенной клетки. Формы апомиксиса зависят от того, из каких частей семязачатка развивается зародыш. При партеногенезе (партенос- девственный) зародыш возникает из неоплодотворенной яйцеклетки. Если зародыш развивается из какой-либо другой клетки гаметофита (антиподы, синергиды), то такой процесс называется апогамия . В случае образования зародыша из клеток нуцеллуса, интегументов, не входящих в гаметофит, говорят об апоспории . Апомиксис распространен среди эволюционно развитых групп растений. За счет апомиксиса растения не зависят от опыляющих агентов
ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ
половой процесс у
покрытосеменных растений, при к-ром оплодотворяются как яйцеклетка, так и
центр. клетка зародышевого мешка.
Д. о. открыл рус. учёный С. Г. Навашин
в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria
orientalis)
. В Д. о. участвуют оба спермия, привносимые в зародышевый
мешок пыльцевой трубкой; ядро одного спермия
сливается с ядром
яйцеклетки,
ядро второго - с полярными ядрами или со вторичным ядром
зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш,
из
центр. клетки - эндосперм.
В зародышевых мешках с трёхклеточным лицевым
аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид,
которая
при этом разрушается (в ней видны остатки ядра синер-гиды и вегетативного ядра
пыльцевой трубки)
; вторая синергида впоследствии отмирает. Далее оба
спермия вместе с изменённой цитоплазмой пыльцевой трубки перемещаются в
щелевидный промежуток между яйцеклеткой и центр. клеткой. Затем спермин
разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром,
другой - проникает в центральную клетку, где контактирует со вторичным
ядром или с одним,а иногда и с обоими полярными ядрами. Спермин теряют свою
цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок;
иногда спермин в виде неизменённых клеток наблюдаются и в
зародышевом мешке.
Двойное оплодотворение; 1 - у рябчика: один из спермиев (а) в контакте с ядром яйцеклетки, второй (б) - с одним из полярных ядер (второе полярное ядро не изображено); 2 - у подсолнечника; а - пыльцевая трубка; 6 - синергиды (одна из них повреждена пыльцевой трубкой); в - яйцеклетка; г - спермий в контакте с ядром яйцеклетки; д - центральная клетка; е - второй спермий в контакте со вторичным ядром зародышевого мешка.
При Д. о. ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние к-рых могут вырьировать. У скерды и нек-рых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у мн. растений они удлинённые, иногда извитые, б. или м. хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Ге-расимова-Навашина) различать два типа Д. о.: премитотическое -ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе) ; постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце к-рой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоид-но. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центр. клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплои-ден. Следствие Д. о. - Ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает неск. пыльцевых трубок, спермий первой из них участвуют в Д. о., спермии остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) С. Г. Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса. Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве Х съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в "Известиях Петербургской Академии Наук".
Мысль о существовании двойного оплодотворения возникла у С. Г. Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии.
Позднее С. Г. Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга - у представителей лютиковых, сложноцветных, орехоцветных, доказав тем самым общность этого явления для всех покрытосеменных.
Открытие С. Г. Навашиным двойного оплодотворения у покрытосеменных сыграло важную роль в науке.
Явление двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксения у кукурузы. Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом.
Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. В ходе этих дискуссий, а главное, благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
Вопросы внутри параграфа: Какие преимущества имеют покрытосеменные перед голосеменными?
Стр. 90. Вопросы и задания после §
1. Что представляет собой гаметофит у разных групп высших растений?
У высших растений половые клетки формируются на гаметофите – гаплоидном поколении, развивающемся из споры. У мхов (сфагнум, кукушкин лен) гаметофит представлен листостебельным растением, у папоротников и семенных растений гаметофит слабо развит и недолговечен. Так, у папоротников, из споры вырастает заросток. Заросток не расчленен на органы, имеет ризоид (примитивные корневые волоски), позднее на нем формируются половые клетки. У моховидных и папоротниковидных половое размножение тесно связано с водой: только при наличии воды сперматозоиды продвигаются к яйцеклетке. У семенных гаметофит сильно редуцирован, например, семенные растения, имеют раздельнополые гаметофиты, которые развиваются из разных спор. Микроспоры дают начало мужскому гаметофиту, мегаспоры – женскому. У цветковых растений мужской гаметофит представлен пыльцевым зерном, созревающим в пыльнике тычинки, и формируются из микроспор. При созревании в пыльцевом зерне образуются две клетки – генеративная и вегетативная. Женский гаметофит, развивающийся из мегаспоры, представляю собой зародышевый мешок, расположенный внутри семязачатка. В нем происходит ряд делений – в результате образуется шесть гаплоидных клеток, среди которых яйцеклетка и одно двойное центральное ядро.
2. Объясните значение терминов «спорофит», «двойное оплодотворение».
Спорофит – это диплоидное поколение высших растений, образующееся при объединении генетических материалов женской и мужской гамет. На спорофите образуются споры. Двойное оплодотворение – это тип оплодотворения высших цветковых растений, в котором участвуют два спермия, один с которых оплодотворяет яйцеклетку, второй - диплоидное центральное яро.
3. Как осуществляется процесс двойного оплодотворения у цветковых растений?
В результате опыления – переноса пыльцы из пыльников на рыльце пестика, пыльцевое зерно – пылинка, начинает прорастать. При участии вегетативной клетки формируется пыльцевая трубка, по которой движется генеративная клетка. Она дает начало двум мужским гаметам спермиям. Спермий – безжгутиковый сперматозоид, не способный к активному движению. Пыльцевая трубка продвигается вниз к завязи и врастает в зародышевый мешок в области пыльцевхода. После этого кончика трубки разрывается и два спермия попадает внутрь зародышевого мешка. Одни из них сливается с яйцеклеткой, образуя зиготу, а другой – с центральным ядром, формируя триплоидное ядро центральной клетки. Из зиготы развивается зародыш с диплоидным набором хромосом, из триплоидной клетки – эндосперм (питательная ткань семени). Из покровов семязачатка закладывается кожура семени, а из стенок завязи – околоплодник. Таким образом, цветок – это укороченный спороносный побег, служащий для образования спор и гамет, для опыления и оплодотворения, после которого образуются плоды и семена.
4. Какую роль играет двойное оплодотворение в размножении цветковых растений?
Благодаря двойному оплодотворению зародыш цветкового растения получает запас питательных веществ, которые расходуют при прорастании. Околоплодник защищает семена и способствует лучшему их прорастанию. Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных, триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов. Увеличение уровня плоидности эндосперма до 3n способствует более быстрому росту ткани по сравнению с диплоидными тканями спорофита
5. Почему гаметофит называют половым поколением, а спорофит – бесполым в жизненном цикле разных групп высших растений?
Гаметофит – это гаплоидная многоклеточная стадия в жизненном цикле растений и водорослей, развивающаяся из спор и производящая половые клетки (гаметы), а споры образуются в результате репродуктивного деления (мейоза) и всегда гаплоидны, то есть содержат одинарный гаплоидный набор хромосом. Спорофит – это диплоидное поколение высших растений, образующееся при объединении генетических материалов женской и мужской гамет.
Двойное оплодотворение
половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка (См. Зародышевый мешок). Д. о. открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия (См. Спермии) сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается Зародыш ,
из центральной клетки - Эндосперм . В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид (См. Синергиды),
которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке. При Д. о. ядра зародышевого мешка находятся в интерфазе (См. Интерфаза) и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать.
У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер. По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа Д. о.: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу (См. Профаза),
в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности (См. Плоидность);
у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие Д. о. - Ксении -
проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в Д. о., спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки. Лит.:
Навашин С. Г., Избр. труды, т. 1, М.- Л., 1951; Магешвар и П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Поддубная Арнольди В. А., Общая эмбриология покрытосеменных растений, М., 1964; Steffen К., Fertilisation, в кн.: Maheshwari P. (ed.). Recent advances in the embryology of angiosperms, Delhi, 1963. И. Д. Романов.
Большая советская энциклопедия. - М.: Советская энциклопедия . 1969-1978 .
Смотреть что такое "Двойное оплодотворение" в других словарях:
Свойственно только цветковым растениям. При двойном оплодотворении один из спермиев сливается с яйцеклеткой, а второй с центральной клеткой зародышевого мешка. Из оплодотворенной яйцеклетки развивается зародыш, из центральной клетки вторичный… … Большой Энциклопедический словарь
Тип полового процесса, свойственный только цветковым растениям. Открыто в 1898 С. Г. Навашиным у лилейных. Д. о. заключается в том, что при формировании семени оплодотворяется не только яйцеклетка, но и центр, ядро зародышевого мешка. Из зиготы… … Биологический энциклопедический словарь
двойное оплодотворение - Тип полового процесса, характерный для цветковых растений: один из спермиев оплодотворяет яйцеклетку, а другой (из той же пыльцевой трубки) оплодотворяет центральное ядро зародышевого мешка, в результате первого процесса образуется диплоидная… … Справочник технического переводчика
Свойственно только цветковым растениям. При двойном оплодотворении один из спермиев сливается с яйцеклеткой, а второй с центральной клеткой зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки вторичный… … Энциклопедический словарь
Double fertilization двойное оплодотворение. Тип полового процесса, характерный для цветковых растений: один из спермиев оплодотворяет яйцеклетку, а другой (из той же пыльцевой трубки
Свойственно только цветковым р ниям. При Д. о. один из спермиев сливается с яйцеклеткой, а второй с центр. клеткой зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центр. клетки вторичный эндосперм семени, содержащий… … Естествознание. Энциклопедический словарь
двойное оплодотворение - процесс оплодотворения, происходящий у покрытосеменных растений, в котором принимают участие оба образующихся спермия. Один из них сливается с яйцеклеткой, второй – с центральной диплоидной клеткой зародышевого мешка. Открыто С. Г. Навашиным в… … Анатомия и морфология растений
ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ - половой процесс у покрытосеменных, заключающийся в слиянии одной мужской гаметы пыльценой трубки (спермия) с яйцеклеткой зародышевого мешка, а второй мужской гаметы с вторичным ядром зародышевого мешка … Словарь ботанических терминов
двойное оплодотворение по навашину - ЭМБРИОЛОГИЯ РАСТЕНИЙ ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ ПО НАВАШИНУ – слияние яйцеклетки и спермия с образованием зиготы (2п) и одновременное слияние другого спермия и двойного ядра с образованием первичного ядра эндосперма (3п). Характерная особенность всех … Общая эмбриология: Терминологический словарь
Двойное оплодотворение , половой процесс у покрытосеменных растений, при котором оплодотворяются как яйцеклетка, так и центральная клетка зародышевого мешка. Двойное оплодотворение открыл русский учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В двойном оплодотворении участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центральной клетки - эндосперм. В зародышевых мешках с трёхклеточным яйцевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синергиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с измененной цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центральной клеткой. Затем спермии разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним, а иногда и с обоими полярными ядрами. Спермии теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермии в виде неизмененных клеток наблюдаются и в зародышевом мешке.
При двойном оплодотворении ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние которых могут вырьировать. У скерды и некоторых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у многих растений они удлинённые, иногда извитые, более или менее хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Герасимова-Навашина) различать два типа двойного оплодотворения: премитотическое - ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце которой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При двойном оплодотворении в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоидно. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центральной клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплоиден. Следствие двойного оплодотворения - ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает несколько пыльцевых трубок, спермий первой из них участвуют в двойном оплодотворении, спермий остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Микроспорогенез протекает в пыльниках тычинок. При этом диплоидные клетки образовательной ткани пыльника в результате мейоза превращаются в 4 гаплоидные микроспоры. Через некоторое время микроспора приступает к митотическому делению и преобразуется в мужской гаметофит - пыльцевое зерно.
Пыльцевое зерно снаружи покрыто двумя оболочками: экзиной и интиной. Экзина - верхняя оболочка более толстая и пропитана спороленнином - жироподобным веществом. Это позволяет пыльце выдерживать существенные температурные и химические воздействия. В экзине находятся проростковые поры, до опыления закрытые «пробочками». Интина содержит целлюлозу и эластична. В пыльцевом зерне имеются две клетки: вегетативная и генеративная.
Мегаспорогенез осуществляется в семязачатке. Из материнской клетки нуцеллуса в результате мейоза образуются 4 мегаспоры, из которых в результате остается только одна. Эта мегаспора сильно разрастается и оттесняет ткани нуцеллуса к интегументам, формируя зародышевый мешок. Ядро зародышевого мешка делится 3 раза митозом. После первого деления два дочерних ядра расходятся к разным полюсам: халазальному и микропилярному , и там делятся два раза. Таким образом, на каждом полюсе находится по четыре ядра. Три ядра у каждого полюса обособляются в отдельные клетки, а два оставшихся перемещаются в центр и сливаются, образуя вторичное диплоидное ядро. На микропилярном полюсе находятся две синергиды и одна более крупная клетка - яйцеклетка . На халазальном полюсе располагаются антиподы . Таким образом, зрелый зародышевыый мешок содержит 7 клеток.
Оплодотворение . Пыльцевые зерна, тем или иным образом попавшие на рыльце, - прорастают. Прорастание пыльцы начинается с разбухания зерна и образования пыльцевой трубки из вегетативной клетки. Пыльцевая трубка прорывает оболочку в более тонком ее месте - так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика. По мере роста пыльцевой трубки в нее переходит ядро вегетативной клетки и генеративной, которая делится и образует два спермия. Через микропиле семязачатка пыльцевая трубка проникает в зародышевый мешок, там она разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, образуя зиготу, которая затем дает начало зародышу семени. Второй спермий сливается с центральным ядром, что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Таким образом, эндосперм у покрытосеменных триплоидный и вторичный, т.к. образуется после оплодотворения.
Весь этот процесс получил название двойного оплодотворения . Он был впервые описан русским ученым С.Г.Навашиным. (1898 г.).
Апомиксис - развитие зародыша из неоплодотворенной клетки. Формы апомиксиса зависят от того, из каких частей семязачатка развивается зародыш. При партеногенезе (партенос- девственный) зародыш возникает из неоплодотворенной яйцеклетки. Если зародыш развивается из какой-либо другой клетки гаметофита (антиподы, синергиды), то такой процесс называется апогамия . В случае образования зародыша из клеток нуцеллуса, интегументов, не входящих в гаметофит, говорят об апоспории . Апомиксис распространен среди эволюционно развитых групп растений. За счет апомиксиса растения не зависят от опыляющих агентов
ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ половой процесс у покрытосеменных растений, при к-ром оплодотворяются как яйцеклетка, так и центр. клетка зародышевого мешка. Д. о. открыл рус. учёный С. Г. Навашин в 1898 на 2 видах растений - лилии (Lilium martagon) и рябчике (Fritillaria orientalis). В Д. о. участвуют оба спермия, привносимые в зародышевый мешок пыльцевой трубкой; ядро одного спермия сливается с ядром яйцеклетки, ядро второго - с полярными ядрами или со вторичным ядром зародышевого мешка. Из оплодотворённой яйцеклетки развивается зародыш, из центр. клетки - эндосперм. В зародышевых мешках с трёхклеточным лицевым аппаратом содержимое пыльцевой трубки обычно изливается в одну из синергид, которая при этом разрушается (в ней видны остатки ядра синер-гиды и вегетативного ядра пыльцевой трубки); вторая синергида впоследствии отмирает. Далее оба спермия вместе с изменённой цитоплазмой пыльцевой трубки перемещаются в щелевидный промежуток между яйцеклеткой и центр. клеткой. Затем спермин разобщаются: один из них проникает в яйцеклетку и вступает в контакт с её ядром, другой - проникает в центральную клетку, где контактирует со вторичным ядром или с одним,а иногда и с обоими полярными ядрами. Спермин теряют свою цитоплазму ещё в пыльцевой трубке или при проникновении в зародышевый мешок; иногда спермин в виде неизменённых клеток наблюдаются и в
При Д. о. ядра зародышевого мешка находятся в интерфазе и обычно значительно крупнее ядер спермиев, форма и состояние к-рых могут вырьировать. У скерды и нек-рых др. сложноцветных ядра спермиев имеют вид двойной скрученной или извитой хроматиновой нити, у мн. растений они удлинённые, иногда извитые, б. или м. хроматизированные, не имеющие ядрышек; обычно спермии представляют собой округлые интерфазные ядра с ядрышками, иногда не отличающиеся по структуре от женских ядер.
По характеру объединения мужских и женских ядер предложено (Е. Н. Ге-расимова-Навашина) различать два типа Д. о.: премитотическое -ядро спермия погружается в женское ядро, хромосомы его деспирализуются; объединение хромосомных наборов обоих ядер происходит в интерфазе (в зиготе); постмитотическое - мужское и женское ядра, сохраняя свои оболочки, вступают в профазу, в конце к-рой начинается их объединение; интерфазные ядра, содержащие хромосомные наборы обоих ядер, образуются лишь после первого митотического деления зиготы. При Д. о. в яйцеклетке сливаются 2 гаплоидных ядра, поэтому ядро зиготы диплоид-но. Число хромосом в ядрах эндосперма зависит от числа полярных ядер в центр. клетке и от их плоидности; у большинства покрытосеменных 2 гаплоидных полярных ядра и эндосперм у них триплои-ден. Следствие Д. о. - Ксении - проявление доминантных признаков эндосперма отцовского растения в эндосперме гибридных семян. Если в зародышевый мешок проникает неск. пыльцевых трубок, спермий первой из них участвуют в Д. о., спермии остальных - дегенерируют. Случаи диспермии, т. е. оплодотворения яйцеклетки двумя спермиями, очень редки.
Рядом точных эмбриологических исследований на различных представителях покрытосеменных (лилейных, лютиковых, сложноцветных) С. Г. Навашин убедительно показал, что эндосперм, как и зародыш, является продуктом полового процесса. Это необычное, свойственное лишь покрытосеменным растениям, явление он назвал двойным оплодотворением. О своем открытии он сообщил в августе 1898 г. на проходившем в Киеве Х съезде русских естествоиспытателей и врачей, а в ноябре того же года опубликовал на эту тему небольшую статью в "Известиях Петербургской Академии Наук".
Мысль о существовании двойного оплодотворения возникла у С. Г. Навашина еще в 1895 г. в процессе работы над изучением халазогамии у грецкого ореха. Окончательное же подтверждение и оформление в стройную теорию эта мысль получила при исследовании оплодотворения у лилии.
Позднее С. Г. Навашин описал двойное оплодотворение и у других цветковых растений, систематически далеко отстоящих друг от друга - у представителей лютиковых, сложноцветных, орехоцветных, доказав тем самым общность этого явления для всех покрытосеменных.
Явление двойного оплодотворения внесло ясность не только в вопрос происхождения эндосперма, но и разъяснило загадочность такого явления, как ксения у кукурузы. Работы Навашина по оплодотворению покрытосеменных растений были встречены ботаниками всего мира с огромным интересом.
Дискуссии о сущности двойного оплодотворения продолжались и в начале ХХ в. В ходе этих дискуссий, а главное, благодаря новым исследованиям, выводы Навашина получили полное подтверждение и дальнейшее развитие.
Водоросли.Общая характеристика. Водоросли – растения, живущие преимущественно в воде. Тело их не расчленено на органы и ткани. Органы размножения одноклеточные. Это одни из древнейших представителей растительного мира. Водоросли бывают одноклеточными (хламидоионада, хлорелла), колониальные (носток) и многоклеточные (спирогира, ламинария,). Цитологические особенности:
Клетки покрыты клеточной стенкой. Характерно наличие хроматофоров , носителей окраски. Хроматофоры – органеллы, в которых происходит фотосинтез. Окраска зависит от глубины обитания водорослей: на большой глубине – буро-красные, ближе к поверхности – зеленые. В хроматофорах всех водорослей содержатся пиреноиды , они синтезируют крахмал.
Одноклеточные формы могут быть подвижными (со жгутиками) и неподвижными.
Также водоросли бывают прикрепленные (бентосные) и свободноплавающие (планктонные) Размножение: Вегетативное – новые водоросли образуются из обрывков нитей, кусков слоевищ и т.д. Бесполое – содержимое одной клетки (зооспорангия) делится многократно, образую новые подвижные клетки (зооспоры). Каждая из них дает начало новой особи. Половое размножение – широко распространено у водорослей. Формы полового процесса разнообразны: изогамия (♂ подвижна, ♀ подвижна, одинаковые по размеру), гетерогамия (♂ подвижна, ♀ подвижна, ♀ больше, чем ♂), оогамия (♂ подвижна, ♀ неподвижна, ♀ больше, чем ♂), конъюгация (сливается содержимое двух вегетативных клеток). Образовавшаяся зигота покрывается толстой клеточной стенкой, накапливает запасные питательные вещества и в состоянии покоя способна легка переносить неблагоприятные условия.
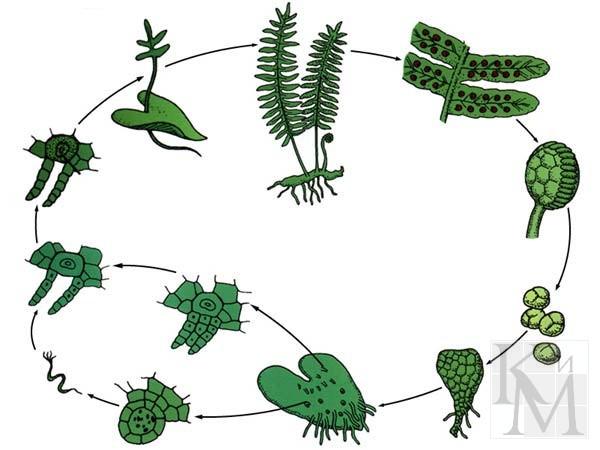
1 .Общая характеристика отдела Папоротниковые . За исключением нескольких родов все папоротники - равноспоровые. У них наблюдается смена поколений с преобладание спорофита над гаметофитом. Спорофит представлен многолетними корневищными травами с крупными, обычно перисто-рассеченными листьями, на нижней стороне которых расположены спорангии. В условия тропического климата встречаются древовидные папоротники. Листья папоротников нарастают верхушкой. То, что у папоротника напоминает лист - вовсе не лист, а по своей природе - целая система ветвей, да ещё расположенных в одной плоскости. Так это и называется - плосковетка, или вайя , или, ещё одно название, - предпобег. В спорангиях в результате мейоза образуются споры (n). Спорангии лопаются, споры высыпаются, прорастает заросток (гаметофит). Заростки представляют собой сердцевидную зеленую пластинку. Точка роста находится в выемке. От нижней стороны отходят ризоиды. На заростке образуются антеридии (в них образуются сперматозоиды) и архегонии (в ней образуется яйцеклетка). Во время дождя или обильной росы сперматозоиды проникают в архегонии и оплодотворяют яйцеклетку. Из зиготы развивается зародыш, затем взрослый спорофит. Современные папоротники насчитывают около 300 родов и 12 000 видов. Основные представители: Щитовник мужской, Страусник обыкновенный, Сальвиния плавающая (разноспоровая)
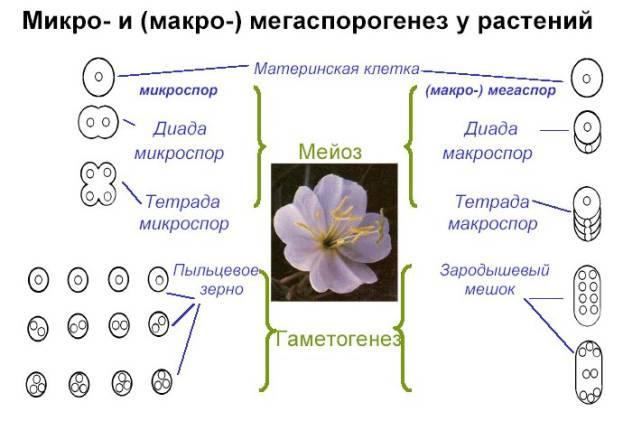
Опыление у покрытосемянных растений. Приспособление цветков к различным видам опыления. Микро- и макроспорогенез у растений Опыление – перенос пыльцы с тычинок на рыльца пестика. Различают самоопыление и перекрестное опыление. Самоопыление: пыльца опыляет рыльце пестика того же цветка (пшеница, ячмень, овес, просо, горох, фасоль, бобы, хлопчатник, лен, томат и др.) . Преобладает у 10% растений. Самоопыление происходит как у раскрывшихся цветков: сельдерейные, так и у закрытых: арахис, фиалка. Однако для эволюционного развития этот вид самоопыления не совершенен, т.к.не несет новых генетических признаков. Поэтому многие растения имеют приспособления, препятствующие самоопылению: Двудомность (♂ и ♀ цветки формируются на разных растениях) Однодомность (♂ и ♀ цветки формируются на одном растении, но в разных цветках) Дихогамия – разновременное созревание пыльцы и рыльца Гетеростилия – рыльца и тычиночные нити различаются по длине Самонесовместимость. Перекрестное опыление : пыльца опыляет рыльца других цветков. Различают 2 типа: гейтоногамия – опыление в пределах одного растения ксеногамия – опыление в пределах разных цветков (наиболее оптимально для эволюционного процесса) Различают несколько видов: Абиотическое – при помощи неживых факторов внешней среды Анемофилия (ветром)
Гидрофилия (водой)
Биотическое – с помощью животных.
Энтомофилия – опыление насекомыми Орнитофилия – опыление мелкими птицами (колибри)
Строение семязачатка. Двойное оплодотворение у покрытосеменных растений.
Строение семязачатка. Семязачаток состоит из центральной части - нуцеллуса и окружающих его одного или двух покровов - интегументов, которые над верхушкой нуцеллуса образуют небольшой канал - микропиле. Семязачаток сообщается с плацентой с помощью семяножки, или фуникулуса. Нуцеллус является аналогом макроспорангия, в котором развивается одна макроспора.Двойное оплодотворение было открыто С.Г.Навашиным в 1898 г. Оплодотворение у покрытосемянных принято называть двойным, т.к. оба спермия сливаются с клетками зародышевого мешка. Один сливается с яйцеклеткой, в результате образуется зигота. Второй сливается с центральным ядром, в результате образуется триплоидная клетка (3n). Прочие клетки зародышевого мешка дегенерируют. После двойного оплодотворения из зиготы развивается зародыш, а из триплойдной клетки – эндосперм (питательная ткань), из нуцеллуса образуется перисперм (дополнительная питательная ткань), из интегумента – семенная кожура, из семязачатка – семя, из завязи – плод. Преимущества двойного оплодотворения заключаются в том, что одновременно с зиготой формируется триплоидная клетка (3n), которая делится быстрее, чем зигота. Соответственно, эндосперм образуется быстрее, чем растет зародыш. Поэтому не нужно запасать питательные вещества до оплодотворения, в отличие от голосеменных, у которых довольно мощный гаплоидный эндосперм развивается до оплодотворения. Этим достигается существенная экономии я энергетических ресурсов организма. Семязачатки покрытосеменных, не обремененные запасающей питательной тканью впрок, развиваются гораздо быстрее, чем у голосеменных.

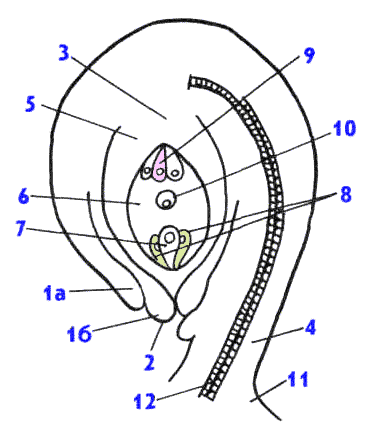
1 - покровы семязачатка, или интегументы (а - наружный, б - внутренний), 2 - микропиле, 3 - халаза, 4 - фуникулус, 5 - нуцеллус, 6 -зародышевый мешок, 7 - яйцеклетка, 8 - синергиды, 9 - антиподы, 10 - вторичное ядро, 11 - плацента, 12 - проводящий пучок. После оплодотворения из зиготы развивается зародыш, из центрального ядра – эндосперм, из нуцеллуса – перисперм, из интигументов – семенная кожура, из семязачатка – семя, из завязи – плод.
Высшие растения. Общая характеристика и цикл развития.
Высшие растения , или Наземные растения ,- тип зелёных растений, которым свойственна дифференциация тканей, в отличие от низших растений - водорослей. К высшим растениям относятся мхи и сосудистые растения (папоротникообразные, псилотовые,хвощевидные, плауновидные, голосеменные и покрытосеменные).
Развитие специализированных тканей было важным условием для выхода растений на сушу. Для комфортного существования в воздушной среде растениям было необходимо развить как минимум эпидермис с устьицами для защиты от высыхания и теплообмена и проводящие ткани для обмена минеральных и органических веществ. Результатом выхода растений на сушу также стало разделение организма растения на корень, стебель и лист.
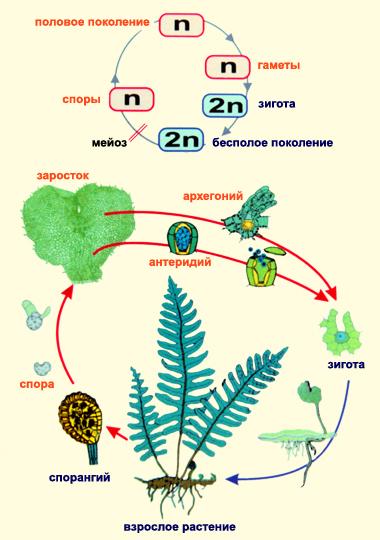
В ЖЦ высших растений происходит чередование полового и бесполого способов размножения и связанное с этим чередование поколений. Бесполое поколение представлено спорофитом (2n), половое – гаметофитом (n). Спорофит – это растение, образующее споры. В многоклеточных спорангиях в результате мейотического деления формируются споры (n). Растения, у которых все споры одинаковые – равноспоровые, у более высокоорганизованных споры разной величины (микроспоры и мегаспоры) – это разноспоровые растения. Гаметофит – растение, образующее гаметы. Гаметы равиваются в многоклеточных органах полового размножения: яйцеклетки – в архегониях, сперматозойды – в антеридиях. Гаметофит вырастает из споры. У равноспоровых растений гаметофит обоеполый, у разноспоровых – однополый. В результате оплодотворения образуется зигота, из которой прорастает новый спорофит. Все высшие растения делятся на 2 группы по доминированию в ЖЦ гаметофита и спорофита: Растения с доминирующим гаметофитом – отдел Моховидные Растения с доминирующим спорофитом – все остальные
В целом для эволюции высших растений характерна тенденция к усложнению и усовершенствованию спорофита при одновременной редукции гаметофита.
Моховидные. Общая характеристика. Цикл развития мха Кукушкин лен. В отдел входит более 25 000 видов сравнительно просто организованных травянистых растений. В цикле развития преобладает гаметофит. У более примитивных форм он представлен талломом, а у остальных – расчленен не стебель и листья. Корней нет, из заменяют ризоиды. Спорофит самостоятельно не существует, развивается на гаметофите, получая от него воду и питательные вещества. Спорофит представляет собой коробочку, где развивается спорангий. Отдел делится на 3 класса: Антоцеротовые, Печеночные, Листостебельные мхи. Наиболее известен вид – Кукушкин лен (Класс Листостебельные мхи). Представляет собой прямостоячие стебли (15-20 см) густо покрытые жесткими острыми листьями. Прикрепляется к земле ризоидами. Гаметофиты раздельнополые. На верхушке мужских особей развиваются антеридии, окруженные красно-бурыми листьями (n), на верхушках женских – архегонии (n). Оплодотворение происходит подвижными двужгутиковыми сперматозоидами во влажную погоду. Из зиготы на верхушке женского гаметофита развивается спорофит (2n), имеющий вид коробочки на длинной ножке. Внутри коробочки – спорангий, где после мейоза образуются споры (n). После образования спор колпачок, а затем крышечка отделяются и споры высыпаются. Из споры сначала образуется протонема, на которой из особых почек формируются листостебельные побеги – гаплоидное поколение (n).
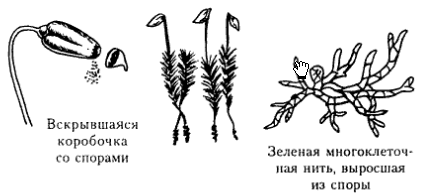

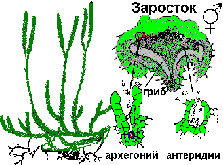
Общая хар-ка отдела Плауновидные. Цикл развития Плауна булавовидного. Плауновидные – очень древняя группа, возникшая в силуре, а в каменноугольном периоде достигла своего расцвета. Это были огромные деревья, образующие целые леса. В современной флоре они представлены вечнозелеными многолетними травами, реже полукустарниками. Около 1000 видов сохранилось. Плауновидные имеют мелкие листья с одной жилкой микрофильного типа. Выделяют 2 класса: равноспоровые Плауновые и разноспоровые Полушниковые. ^ Плаун булавовидный . В цикле развития преобладает спорофит (2n). Он представляет собой длинные стелющиеся ветвящиеся побеги густо усажены жесткими мелкими листьями. От стебля отходят тонкие придаточные корни. В середине лета на верхушках появляются спороносные колоски. Колосок состоит из оси и сидящих на ней листочков (спорофиллов). На верхней стороне спорофиллов находится спорангий на короткой ножке. В результате мейоза в нем образуются споры (n). Из спор развивается заросток (гаметофит) (n). Заросток развивается под землей. От нижней его части отходят ризоиды. Через них он врастает в грибы, образуя микоризу. Живет в симбиозе с грибом, питается от него. Растет очень долго (15-20 лет). На верхней стороне его образуется антеридий и архегонии. Двужгутиковый сперматозоид выходи из антеридия, проникает в архегоний и оплодотворяет яйцеклетку. В результате образуется зигота, из которой развивается новый спорофит.

Голосеменные – это древний отдел растений, которые характеризуются наличием семязачатков с последующим развитием семени, но не образуют цветков и плодов. К голосеменным относятся около 800 современных видов. Преобладающее большинство видов этого отдела (более 500) составляют класс хвойных растений.
Типичный
представитель – СОСНА ОБЫКНОВЕННАЯ
Сосна
– разноспоровое однодомное растение.
Микро- и мегаспорангии образуются на
одном и том же дереве в отдельных
шишках.
Женские
шишки состоят из оси и семенных чешуй,
на верхней стороне которых развивается
по 2 семязачатка. Семязачаток состоит
из нуцеллуса, покрытого интегументом,
в котором имеется отверстие – микропиле.
Внутри нуцеллуса путем мейоза образуются
4 мегаспоры, 3 из них отмирают, а одна
прорастает в женский гаметофит –
зародышевый мешок.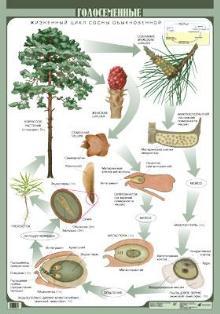
Мужские шишки собраны в сошишие на верхушках удлиненных побегов. В микроспорангиях в результате мейоза из клеток спорогенной ткани возникают микроспоры (n). Микроспора покрыта 2 оболочками: интина (внутренняя) и экзима (внешняя). Экзима отстает от интины, образую 2 воздушных мешка. Увеличивая поверхность пыльцы, они способствуют переносу ее ветром, а при попадании пылинки в жидкость выполняют роль поплавков. Мужские шишки после производства пыльцы быстро засыхают и опадают с ветвей. Пыльцевое зерно содержит мужской гаметофит, состоящий из двух клеток – вегетативной и генеративной. Опыление происходит с помощью ветра в начале лета. Пыльца прилипает к капле клейкой жидкости, которая выделяется в области микропиле, вегетативная клетка начинает удлиняться в пыльцевую трубку, чешуи женских шишек смыкаются.
В начале весны происходит оплодотворение: генеративная клетка делится с образованием двух спермиев, которые движутся по пыльцевой трубке. Далее один из спермиев сливается с яйцеклеткой, а другой отмирает. Из оплодотворенной яйцеклетки (зиготы) формируется зародыш семени, а семязачаток превращается в семя.
Семена у сосны созревают на второй год, высыпаются из шишек и, подхваченные животными или ветром, переносятся на значительные расстояния.
Классификация покрытосеменных растений. Отдел Покрытосеменные делится на 2 класса: однодольный и двудольные. Эволюция среди этих классов шла разными путями, вследствие чего каждый из них имеет характерные отличия. Эти отличия настолько сильны, что двудольные не скрещиваются с однодольными и прививок между ними не получено. Основные различия
|
Класс Двудольные |
Класс Однодольные |
|
Около 190 000 видов |
Около 63 000 видов |
|
Семя |
|
|
Зародыш с двумя семядолями |
Зародыш с одной семядолей |
|
Семядоли латеральные (боковые), обычно с тремя главными проводящими пучками |
Семядоля терминальная (верхушечная), с двумя проводящими пучками |
|
Корень |
|
|
Система главного корня, стержневая |
Придаточная, мочковатая КС |
|
Вторичное утолщение (камбий есть) | |
|
Стебель |
|
|
Сплошное расположение проводящих тканей или пучки, расположенные по кругу |
Разбросанное расположение проводящих пучков |
|
Вторичное утолщение, камбий есть |
Отсутствие вторичного утолщения (камбия нет) |
|
Лист |
|
|
Простой, сложный, более или менее четко разделен на черешок и пластинку |
Простой, обычно не расчлененный на черешок и пластинку |
|
Жилкование перистое или пальчатое |
Жилкование параллельное или дуговое |
|
Цветок |
|
|
Пяти-, много-, четырехчленный |
Трехчленный |
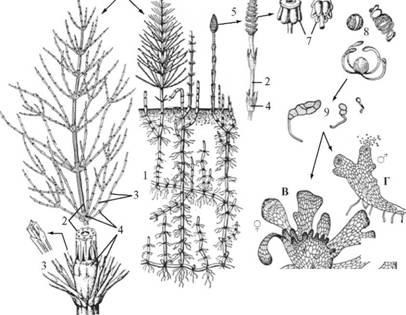
Отдел Хвощевидные. Общая характеристика. Цикл развития Хвоща полевого. Отдел Хвощевидные в настоящее время представлены только одним родом Хвощ и 25 видами. В цикле развития преобладает спорофит (2n), который представлен в виде многолетней травы. От корневищ отходят придаточные корни и надземные побеги, обычно однолетние. Побеги имеют типичное членистое строение. От узлов отходят бурые чешуйчатые листья, сросшихся в трубчатое влагалище. Функцию фотосинтеза выполняют стебли. Споры (n) образуются в спороносных колосках, развивающихся в зависимости от вида хвоща на верхушках ассимилирующих или на специализированных спороносных бесхлорофилльных побегах. На оси колосков находятся спорангиофоры, имеющие вид щитка на ножке, к которому прикреплены 8-10 спорангиев. Образовавшиеся после мейоза споры покрыты толстой оболочкой, снабжены лентовидными пружинками – элатерами (при высыхании они раскручиваются, при увлажнении закручиваются, благодаря чему сцепляются между собой). В результате прорастания спор образуются мужские и обоеполые заростки. Заростки очень маленькие, имеют вид рассеченных пластинок, от нижней поверхности которых отходят ризоиды. Гаметофит созревает в течение 3-5 недель. На концах лопастей мужских заростков развиваются антеридии, в средней части архегонии. Архегонии развиваются раньше, чтобы увеличить вероятность оплодотворения чужими сперматозоидами. Сперматозоиды много жгутиковые. Из зиготы развивается новый спорофит.

Строение
цветка, особенности строения околоцветника
.
Цветок
– сложный репродуктивный орган
покрытосеменных растений.
Цветок
имеет ось (цветоложе), несущее листочки
околоцветника, тычинки и пестик (или
пестики).
Если
цветоножка не выражена, цветок называют
сидячим.
Части
цветка делятся на фертильные
(тычинка, пестик) и стерильные (чашечка,
венчик, околоцветник).
Цветок,
содержащий тычинки и пестики.
называют обоеполым
(у
70% цветков). Но бывают случаи, когда
цветки однополые
.
Растения с однополыми цветками,
находящимися на одном и том же экземпляре,
называют однодомными
(кукуруза,
дуб, бук, огурец, тыква), если цветки
расположены на разных экземплярах
– двудомные
(конопля,
ива, тополь).
В
отличие от симметрии различают:
правильные
(актиноморфные) цветки (если можно
провести несколько осей симметрии) –
капустные, гвоздичные, розовые
неправильные
(зигоморфные)
цветки (если можно провести только одну
ось) – бобовые, яснотковые
нессиметричные
(ассиметричные) – если нельзя провести
даже одну ось симметрии – валериана
лекарственная.
Околоцветник
– это стерильная часть цветка, является
покровом, защищающим более нежные
тычинки и пестики. Он бывает двойной и
простой.
^
Двойной
околоцветник
дифференцирован
на чашечку и венчик.
Чашечка
состоит из чашелистиков, образует
наружный круг околоцветника. Чашелистики
обычно имеют небольшие размеры и зеленую
окраску. Главная функция – защита цветка
до раскрывания бутона.
Венчик
состоит из лепестков. Лепестки имеют
более крупные размеры и различаются по
окраске. Окраску определяют различные
пигменты:
антоциан
(розовая, красная, синяя, фиолетовая),
каротиноиды (желтая, оранжевая, красная),
антохлор (лимонно-желтая), антофеин
(коричневая)
Аромат
цветка создается эфирными маслами.
Венчик
может быть раздельнолепестным (лютик,
земляника) и сростнолепестным (картофель,
шалфей).
^
Простой
околоцветник
не
дифференцирован на чашечку и венчик,
состоит из совокупности однородных
листков околоцветника. Может быть
венчиковидным (тюльпан) и чашечковидным
(манжетка) 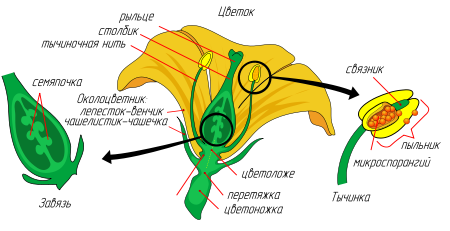
Простые и двойные околоцветники бывают раздельнолепестными (яблоня, тюльпан) и сростнолепестными (колокольчик, ландыш). Кроме чашечки и венчика с стерильным частям цветка относят еще нектарник . ФОРМУЛА И ДИАГРАММА ЦВЕТКА Строение цветка можно выразить в виде формулы . При ее составлении используют следующие обозначения:
Цветок правильный (актиноморфный) - цветок неправильный (зигоморфный) Ca- чашечка(calyx) Co- венчик(corolla) P - простой околоцветник(perigonium) A - тычинки(андроцей) (androceum) G - пестик, плодолистики (гинецей) (gynoeceum)
Рядом с буквенными выражениями частей цветка цифрами указывается количество элементов (пятилепестный венчик - Co 5 , шестичленный андроцей - A 6), а в том случае если их число в цветках одного и того же вида непостоянно (обычно больше 12)- символом . Если элементы цветка сросшиеся, то их число заключается в скобки: сросшийся пятичленный венчик - Co (5) , двубратственный андроцей - A (9,1) . Если элементы цветка расположены кругами, то между количеством элементов в каждом круге ставится знак «+» (P 3+3). Положение завязи в цветке обозначается чёрточкой. При верхней завязи цветок является подпестечным, поэтому под цифрой числа плодолистиков ставят чёрточку G () , при обозначении нижней завязи - цветок надпестичный - чёрточку ставят над цифрой G () . При простом околоцветнике знаки чашечки и венчика не применяются, и он обозначается буквой P (перигониум).. Например, формула цветка лилии и тюльпана: P 3+3 A 3+3 G ()
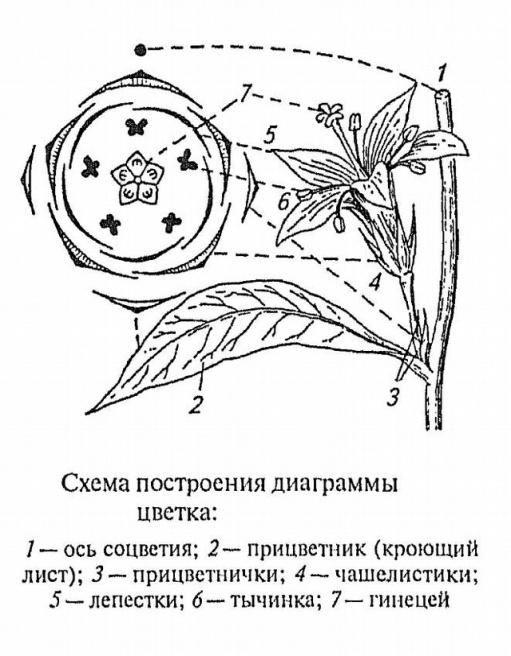
Диаграммой
называют схематическую проекцию цветка
на плоскости в следующей ориентации:
ось соцветия вверху, кроющий лист –
внизу.
Классификации плодов. Морфогенетическая классификация основана на типе гинецея: Искусственная классификация плодов основана главным образом на признаках внешней морфологии: Простой плод развивается только из завязи одного пестика (монокарпный, ценокарпный гинецей). Сборный плод (сложный плод) формируется из завязей нескольких свободных пестиков (апокарпный гинецей). Соплодие – сросшиеся в единое целое несколько или много плодов, образовавшихся из цветков одного соцветия. В зависимости от консистенции околоплодника различают: сухие и сочные В зависимости от числа семян различают: многосемянные и односемянные. В зависимости от вскрывания околоплодника различают: невскрывающиеся и вскрывающиеся плоды.
Сочные плоды. В эволюции сочные плоды появились как производные сухих. Стенка СП может развиваться либо из стенки завязи (перикарпа), либо из стенки завязи, сросшейся с цветочной трубкой или с вогнутым цветоложем. Стенка незрелого СП имеет плотную структуру, по мере созревания плода она становится мягче. Созревание сопровождается как правило изменением окраски: за счет разрушения хлорофилла и накопления либо каротиноидов (желтый, оранжевый цвета), либо антоцианов (красные, синие). Созревание также сопровождается накоплением углеводов. Среди сочных плодов выделяют многосемянные сочные плоды Гранатина - развивается из нижней многогнездной завязи с двумя этажами. Мякоть образуется из сочного наружного слоя семенной кожуры. Околоплодник и ткани цветочной трубки подсыхают, образую твердую кожуру Геспередий (померанец) – цитрусовые, формируется из синкарпного гинецея. Экзокарп – желтая ткань, покрыта слоем кутикулы, воска и маслянистыми железками. Мезокарп – белая ткань. Эндокарп – пленчатый. Субэпидермальные клетки эндокарпа формируют соковые мешочки (мякоть плода). Тыквина – экзокарп жесткий, одревесневающий, кожистый; мякоть образована мезо- и эндокарпом (тыква, дыня) или разросшимися плацентами (арбуз). Образуется из паракарпного гинецея. Яблоко – формируется из апокарпного гинецея
Ягода – сочный мясистый эндо и мезокарпий, тонкий пленчатый экзокарпий Односемянные сочные плоды: Костянка – невскрывающийся монокарпий с мясистым сочным съедобным мезокарпом и склерифицированным эндокарпом (косточка). (слива, вишня,черешня, абрикос, персик, алыча).
Простые сухие плоды:
|
Многосемянные |
Односемянные |
|
|
Вскрывающиеся |
Распадающиеся |
|
|
Листовка (рогатый василек) |
Двусемянка (морковь, укроп) |
Зерновка (рожь, пшеница) |
|
Боб (Бобовые) |
Двукрылатка (клен) |
Семянка (подсолнечник) |
|
Стручок (капуста, сурепица) |
Четырехорешек |
Крылатка (ясень, вяз, береза) |
|
Стручочек (пастушья сумка) |
Дробная коробочка |
Орех (фундук) |
|
Коробочка (мак, белена, хлопчатник) |
Членистый боб (копеечник, вязель) |
Орешек (липа, гречиха) |
|
Членистый стручок (редька дикая) |
Желудь (дуб, бук) |
|
Общая характеристика высших растений, чередование гаплоидной и диплоидной фаз у высших растений. Высшие растения – одно из подцарств царства Растений, объединяющее не менее 300 000 видов – от мхов до покрытосеменных. Их происхождение связано с выходом на сушу морских многоклеточных водорослей. В процессе приспособления к наземным условиям существования возникли новые типы растений с новыми признаками: 1. На смену таллому у высших растений формируется тело, расчлененное на органы: корень и побег (листья и стебель) 2. Усложняется внутреннее строение, появляются специализированные ткани: покровные (защита растений), проводящие, механические (опора) 3. Формируются многоклеточные органы размножения, т.к. многоклеточные стенки надежнее защищают гаметы или споры 4. Зигота развивается в типичный многоклеточный зародыш 5. Задержка зиготы внутри архегония – еще одна функция защиты в условиях суши 6. У высокоорганизованных семенных растений приспособление к наземному образу жизни выразилось в полной независимости полового размножения от капельно-жидкой воды 7. У низших растений встречаются различные варианты полового процесса: изогамия (мужская и женская половая клетка одного размера подвижные), гетерогамия (женская половая клетка больше, чем мужская, обе подвижные), оогамия (женская половая клетка неподвижная, больше, чем мужская). У высших растений встречается только оогамия. 8. У высших растений у цикле развития всегда происходит чередование поколений (спорофит – бесполое поколение, гаметофит – половое поколение). На спорофите (2n) за счет мейотического деления образуются гаплоидные споры (n). Из спор развивается гаплоидный гаметофит (n), производящий гаплоидные гаметы. При их слиянии в зиготе восстанавливается набор хромосом. Из зиготы вновь развивается диплоидный спорофит (2n) Высшие растения разделяются на отделы.


