Серая жаба прямое или непрямое развитие. Этапы развития человека и животных
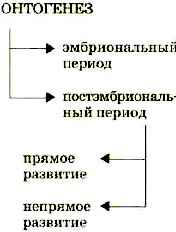
1. Прямое развитие - развитие, при котором появившийся организм идентичен по строению взрослому организму, но имеет меньшие размеры и не обладает половой зрелостью. Дальнейшее развитие связано с увеличением размеров и приобретением половой зрелости. Например: развитие рептилий, птиц, млекопитающих.
2. Непрямое развитие (личиночное развитие, развитие с метаморфозом) - появившийся организм отличается по строению от взрослого организма, обычно устроен проще, может иметь специфические органы, такой зародыш называется личинкой. Личинка питается, растет и со временем личиночные органы заменяются органами, свойственными взрослому организму (имаго). Например: развитие лягушки, некоторых насекомых, различных червей.Постэмбриональное развитие сопровождается ростом.
Билет №26. Биосоциальная природа человека как отражение эволюционно обусловленной иерархии живой природы. Значение биологического наследия человека в современных условиях жизни.
На планете среди других существ людям принадлежит уникальное место, что обусловлено приобретением ими в процессе антропогенеза особого качества - социальной сущности. Это означает, что уже не биологические механизмы, а в первую очередь общественное устройство, интеллект, производство, труд обеспечивают выживание, всесветное и даже космическое расселение, благополучие человечества. Социальность, однако, не противопоставляет людей остальной живой природе. Человек остается включенным в систему органического мира. Этот мир складывался и развивался на протяжении большей части истории планеты независимо от человеческого фактора, более того, на определенном этапе своего развития он этот фактор породил. Человечество составляет своеобразный, но неотъемлемый компонент биосферы. Крупный отечественный патолог И. В. Давыдовский писал, что естественность и законность болезней вытекают из основных свойств жизни, а именно из универсального и важнейшего свойства организмов - приспосабливаться к меняющимся условиям внешней среды. По его мнению, полнота такого приспособления и есть полнота здоровья.
Билет №27 Гаметогенез или предзародышевое развитие - процесс созревания половых клеток, или гамет. Поскольку в ходе гаметогенеза специализация яйцеклеток и спермиев происходит в разных направлениях, обычно выделяют овогенез и сперматогенез соответственно. Гаметогенез закономерно присутствует в жизненном цикле ряда простейших, водорослей, грибов, споровых и голосемянных растений, а также многоклеточных животных. В некоторых группах гаметы вторично редуцированы (сумчатые и базидиевые грибы, цветковые растения). Наиболее подробно процессы гаметогенеза изучены у многоклеточных животных.
Гаметогенез
(от гаметы и греч. genesis - происхождение), процесс развития и формирования половых клеток - гамет. Г. мужских гамет (сперматозоидов, спермиев) называют сперматогенезом, женских гамет (яйцеклеток) - оогенезом. У животных и растений Г. протекает различно, в зависимости от места мейоза в жизненном цикле этих организмов.
У многоклеточных животных Г. происходит в специальных органах - половых железах, или гонадах (яичниках, семенниках, гермафродитных половых железах), и складывается из трёх основных этапов: 1) размножение первичных половых клеток - гаметогониев (сперматогониев и оогониев) путём ряда последовательных митозов, 2) рост и созревание этих клеток, называют теперь гаметоцитами (сперматоцитами и ооцитами), которые, как и гаметогонии, обладают полным (большей частью диплоидным) набором хромосом. В это время совершается основное событие Г. у животных - деление гаметоцитов путём мейоза, приводящее к редукции (уменьшению вдвое) числа хромосом в этих клетках и превращению их в гаплоидные клетки (см. Гаплоид) - сперматиды и оотиды; 3) формирование сперматозоидов (либо спермиев) и яйцеклеток; при этом яйцеклетки одеваются рядом зародышевых оболочек, а сперматозоиды приобретают жгутики, обеспечивающие их подвижность. У самок многих видов животных мейоз и формирование яйца завершаются после проникновения сперматозоида в цитоплазму ооцита, но до слияния ядер сперматозоида и яйцеклетки.
У растений Г. отделен от мейоза и начинается в гаплоидных клетках - в спорах (у высших растений - микроспоры и мегаспоры). Из спор развивается половое поколение растения - гаплоидный гаметофит, в половых органах которого - гаметангиях (мужских - антеридиях, женских - архегониях) путём митозов происходит Г. Исключение составляют голосеменные и покрытосеменные растения, у которых сперматогенез идёт непосредственно в прорастающей микроспоре - пыльцевой клетке. У всех низших и высших споровых растений Г. в антеридиях - это многократное деление клеток, в результате которого образуется большое число мелких подвижных сперматозоидов. Г. в архегониях - формирование одной, двух или нескольких яйцеклеток. У голосеменных и покрытосеменных растений мужской Г. состоит из деления (путём митоза) ядра пыльцевой клетки на генеративное и вегетативное и дальнейшего деления (также путём митоза) генеративного ядра на два спермия. Это деление происходит в прорастающей пыльцевой трубке. Женский Г. у покрытосеменных растений - обособление путём митоза одной яйцеклетки внутри 8-ядерного зародышевого мешка. Основное различие Г. у животных и растений: у животных он совмещает в себе превращение клеток из диплоидных в гаплоидные и формирование гаплоидных гамет; у растений Г. сводится к формированию гамет из гаплоидных клеток.
Гаметы (от греч. ?????? - жена, ??????? - муж) - репродуктивные клетки, имеющие гаплоидный (одинарный) набор хромосом и участвующие в гаметном, в частности, половом размножении. При слиянии двух гамет в половом процессе образуется зигота, развивающаяся в особь (или группу особей) с наследственными признаками обоих родительских организмов, продуцировавших гаметы.
У некоторых видов возможно и развитие в организм одиночной гаметы (неоплодотворённой яйцеклетки) - партеногенез.
Морфология гамет и типы гаметогамии
Морфология гамет различных видов достаточно разнообразна, при этом продуцируемые гаметы могут отличаться как по хромосомному набору (при гетерогаметности вида), величине и подвижности (способности к самостоятельному передвижению), при этом гаметный диморфизм у различных видов варьирует в широких пределах - от отсутствия диморфизма в виде изогамии до своего крайнего проявления в форме оогамии.
Изогамия
Если сливающиеся гаметы морфологически не отличаются друг от друга величиной, строением и хромосомным набором, то их называют изогаметами, или бесполыми гаметами. Такие гаметы подвижны, могут нести жгутики или быть амёбовидными. Изогамия типична для многих водорослей.
Анизогамия (гетерогамия)
Гаметы, способные к слиянию, различаются по размерам, подвижные микрогаметы несут жгутики, макрогаметы могут быть как подвижны (многие водоросли), так и неподвижны (лишённые жгутиков макрогаметы многих протистов).
Способные к слиянию гаметы одного биологического вида резко различаются по размерам и подвижности на два типа: малые подвижные мужские гаметы - сперматозоиды - и крупные неподвижные женские гаметы - яйцеклетки. Различие размера гамет обусловлено тем, что яйцеклетки содержат запас питательных веществ, достаточный для обеспечения нескольких первых делений зиготы при её развитии в зародыш.
Мужские гаметы - сперматозоиды - животных и многих растений подвижны и обычно несут один или несколько жгутиков, исключением являются лишённные жгутиков мужские гаметы семенных растений - спермии, которые доставляются к яйцеклетке при прорастании пыльцевой трубки, а также безжгутиковые сперматозоиды (спермии) нематод и членистоногих.
Хотя сперматозоиды несут митохондрии, при оогамии от мужской гаметы к зиготе переходит только ядерная ДНК, митохондриальная ДНК (а в случае растений и пластидная ДНК) обычно наследуется зиготой только от яйцеклетки.
Билет №28 . Нетрадиционное наследование (геномный импринтинг, однородительская дисомия, экспансия тринуклеотидных повторов, митохондриальное наследование).
ЭКСПАНСИЯ ТРИНУКЛЕОТИДНЫХ ФРАГМЕНТОВ - патологическое состояние: вариант генетической мутации, характеризующийся появлением в ДНК "бессмысленных" повторов тринуклеотидов, которые могут приводить к дезорганизации функционирования ДНК или синтезу патологического белка, накапливающегося в клетках, что приводит к гибели клетки. Лежит в основе ряда заболеваний (болезни Гентингтона, болезни Кеннеди, спиноцеребеллярных дегенерации и т.д.), тяжесть которых зависит от числа повторов тринуклеотидов. Общая особенность этой группы заболеваний - более раннее начало и нарастание тяжести их клинических проявлений из поколения в поколение, что обычно отражает увеличение числа тринуклеотидных повторов (феномен антиципации).В последнее время выделяется еще один тип наследования - митохондриальный. Митохондрии передаются с цитоплазмой яйцеклеток. Спермии не имеют митохондрий, поскольку цитоплазма элиминируется в процессе созревания мужских половых клеток. В яйцеклетке содержится около 25000 митохондрий. Каждая митохондрия содержит кольцевую хромосому. Генные мутации в митохондриальной ДНК обнаружены при атрофии зрительного нерва Лебераф, митохондриальных миопатиях, прогрессирующих офтальмоплегиях. Болезни, обусловленные данным типом наследственности, передаются от матери и дочерям, и сыновьям в равной степени. Больные отцы болезнь не передают ни дочерям, ни сыновьям.
Геномный импринтинг - эпигенетический процесс, при котором экспрессия определенных генов осуществляется в зависимости оттого, от какого родителя поступил аллель гена. Это ненаследуемый процесс, который не подчиняется наследованию по Менделю. Импринтинг генов вызывает экспрессию аллелей гена полученных от матери в случае генов H19 или CDKN1C и от отца в случае гена IGF2. Импринтинг некоторых генов в составе генома показан для насекомых, млекопитающих и цветковых растений.
Импринтинг генов осуществляется с помощью процесса метилирования ДНК. Если по каким-то причинам импринтинг не сработает, это может привести к появлению генетических нарушений (например Синдром Прадера-Вилли).
Однородительская дисомия, то есть наследование обеих копий целой хромосомы или ее части от одного родителя (при отсутствии соответствующего генетического материала от другого родителя), является исключением из менделевских принципов наследования. Она встречается редко и вызывает, например, синдром Прадера-Вилли и синдром Ангельмана.
Роль дисомии в патологии во многом усугубляется геномным импринтингом, который приводит к неодинаковой экспрессии материнской и отцовской копий гена.
Возможный механизм дисомии - элиминация лишней хромосомы у плода с трисомией на ранних стадиях эмбриогенеза. Болезнь проявляется в том случае, если элиминируется лишняя хромосома, происходящая из нормальной гаметы.
Однородительская дисомия была описана при муковисцидозе, когда оба мутантных аллеля наследовались от одного родителя. В таких случаях дисомия имитирует аутосомно-рецессивное наследование.
У 20-30% больных с синдромом Прадера-Вилли, имеющих по данным цитогенетического исследования нормальный кариотип, с помощью молекулярно-биологических методов обнаруживается дисомия материнской 15-й хромосомы. Отцовская 15-я хромосома у таких больных отсутствует.
Предполагают, что однородительская дисомия является причиной внутриутробной задержки развития, умственной отсталости и микроцефалии. Эти предположения пока не подтверждены молекулярно-биологическими исследованиями.
Билет №29 СМ. №2
Билет №30
Феноти?п - (от греческого слова phainotip - являю, обнаруживаю) совокупность характеристик, присущих индивиду на определённой стадии развития. Фенотип формируется на основе генотипа, опосредованного рядом внешнесредовых факторов. У диплоидных организмов в фенотипе проявляются доминантные гены.
Фенотип - совокупность внешних и внутренних признаков организма, приобретённых в результате онтогенеза (индивидуальное развитие)
Несмотря на кажущееся строгое определение, концепция фенотипа имеет некоторые неопределенности. Во-первых, большинство молекул и структур кодируемых генетическим материалом, не заметны во внешнем виде организма, хотя являются частью фенотипа. Например, именно так обстоит дело с группами крови человека. Поэтому расширенное определение фенотипа должно включать характеристики, которые могут быть обнаружены техническими, медицинскими или диагностическими процедурами. Дальнейшее, более радикальное расширение может включать приобретенное поведение или даже влияние организма на окружающую среду и другие организмы. Например, согласно Ричарду Докинзу, плотину бобров также как и их резцы можно считать фенотипом генов бобра.
Фенотип можно определить как «вынос» генетической информации навстречу факторам среды. В первом приближении можно говорить о двух характеристиках фенотипа: а) число направлений выноса характеризует число факторов среды, к которым чувствителен фенотип, - мерность фенотипа; б) «дальность» выноса характеризует степень чувствительности фенотипа к данному фактору среды. В совокупности эти характеристики определяют богатство и развитость фенотипа. Чем многомернее фенотип и чем он чувствительнее, чем дальше фенотип от генотипа, тем он богаче. Если сравнить вирус, бактерию, аскариду, лягушку и человека, то богатство фенотипа в этом ряду растет.
Геном - вся совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Он обеспечивает формирование видовых характеристик организмов в ходе их онтогенеза. Генотип - совокупность генов, образованная при половом размножении в процессе оплодотворения при объединении геномов двух родительских клеток, генетическая конституция организма, представляющая собой совокупность всех наследственных задатков его клеток, заключенных в их хромосомном наборе - кариотипе. Фенотип - видовые и индивидуальные морфологические, физиологические и биохимические свойства на всем протяжении индивидуального развития. Ведущая роль в формировании фенотипа - наследственная информация, заключенная в генотипе. Наряду с этим результат наследственной программы (в генотипе) зависит от условий, в которых осуществляется этот процесс. В случае гетерозиготности развитие данного признака будет зависеть от взаимодействия аллельных генов. Доминирование - это такое взаимодействие аллельных генов, при котором проявление одного из аллелей (А) не зависит от присутствия в генотипе другого (А’). Этот аллель доминантный, второй рецессивный (пример: группа крови). Неполное доминирование - фенотип гетерозигот ВВ’ отличается от фенотипа гомозигот по обеим аллелям (ВВ, В’В’) промежуточным проявлением признака. Это происходит, т.к. аллель, способная сформировать нормальный признак находится у гетерозигот в двойной дозе ВВ, а у гомозигот ВВ’. Генотипы отличаются экспрессивностью (степень выраженности признака). Пример: заболевания у человека, проявляющиеся клинически у гетерозигот, а у гомозигот заканчивающиеся смертью. Кодоминирование - каждый из аллелей проявляет свое действие, в результате - промежуточный вариант признака (Группа крови, аллели которые по отдельности формируют 2 и 3 группы крови, вместе образуют 4). Аллельное исключение - вид взаимодействия аллельных генов в генотипе. Например, инактивация одного из аллелей в сосотаве Х-хромосомы способствует тому, что разных клетках организма, мозаичных по функционирующей хромосоме, фенотипически проявляются разные аллели.
Билет №31 . Генетический гомеостаз и механизмы его обеспечения на разных уровнях организации жизни.
ГОМЕОСТАЗ генетический - способность популяции поддерживать динамическое равновесие генетического состава, что обеспечивает ее жизнеспособность.Гомеостатические системы обладают следующими свойствами:
Нестабильность системы: тестирует, каким образом ей лучше приспособиться.Стремление к равновесию: вся внутренняя, структурная и функциональная организация систем способствует сохранению баланса.Непредсказуемость: результирующий эффект от определённого действия зачастую может отличаться от того, который ожидался.
Механизмы гомеостаза: обратная связь.Когда происходит изменение в переменных, наблюдаются два основных типа обратной связи, или фидбэка, на которые реагирует система:Отрицательная обратная связь, выражающаяся в реакции, при которой система отвечает так, чтобы изменить направление изменения на противоположное. Так как обратная связь служит сохранению постоянства системы, это позволяет соблюдать гомеостаз. Например, когда концентрация углекислого газа в организме человека увеличивается, лёгким приходит сигнал к увеличению их активности и выдыханию большего количество углекислого газа.Терморегуляция - другой пример отрицательной обратной связи. Когда температура тела повышается (или понижается) терморецепторы в коже и гипоталамусе регистрируют изменение, вызывая сигнал из мозга. Данный сигнал, в свою очередь, вызывает ответ - понижение температуры (или повышение).Положительная обратная связь, которая выражается в усилении изменения переменной. Она оказывает дестабилизирующий эффект, поэтому не приводит к гомеостазу. Положительная обратная связь реже встречается в естественных системах, но также имеет своё применение. Например, в нервах пороговый электрический потенциал вызывает генерацию намного большего потенциала действия. Свёртывание крови и события при рождении можно привести в качестве других примеров положительной обратной связи.Устойчивым системам необходимы комбинации из обоих типов обратной связи. Тогда как отрицательная обратная связь позволяет вернуться к гомеостатическому состоянию, положительная обратная связь используется для перехода к совершенно новому (и, вполне может быть, менее желанному) состоянию гомеостаза, - такая ситуация называется «метастабильность». Такие катастрофические изменения могут происходить, например, с увеличением питательных веществ в реках с прозрачной водой, что приводит к гомеостатическому состоянию высокой эвтрофикации (зарастание русла водорослями) и замутнению.
Билет №32 Половой процесс и эволюция его форм.характеристики гамет. Препарат-зигота. Задача на гемофилию
Билет №33 Транскрипция.Процессинг и сплайсинг.Альтернативный сплайсинг
Билет №34, 47. Биологические ритмы и факторы внешней среды. Хронобиология и хрономедицина, понятие о десинхронозах.
Хронобиология (от «Chrono», «Chronos» - «время») - область науки, которая исследует периодические (циклические) феномены, протекающие у живых организмов во времени, и их адаптацию к солнечным и лунным ритмам . Эти циклы именуют биологические ритмы (БР).Хронобиологические исследования включают, но не ограничиваются ими, работы в области сравнительной анатомии, физиологии, генетики, молекулярной биологии и биологии поведения организмов . Другие аспекты включают исследование развития, воспроизведения, экологии и эволюции видов.
Описание.Синхронизация уровня и длительности биологической активности с внешними факторами у живых организмов происходят при многих существенных биологических процессах. Это происходит
· у животных (еда, сон, спаривание, зимовка, миграция, клеточная регенерация, и т. д.),
· у растений (движения листа, фотосинтез и т. д.).
Наиболее важный ритм в хронобиологии - суточный ритм, примерно 24-часовой цикл физиологических процессов у растений и животных. (Слово «циркадный» приходит из латыни, - «цирка» означает «около», «примерно», и «диас» - «день», «сутки», то есть «циркадианный» или «циркадный» - это «околосуточный»).
Есть и другие важные циклы:
· инфрадианные, более долгосрочные, такие как ежегодные циклы миграции или воспроизведства, выявленные у некоторых животных, или человеческий менструальный цикл.
· ультрадианные ритмы, краткие циклы, такие как 90-минутный цикл REM-сна у людей, 4-часовой назальный цикл или 3-часовой цикл продуцирования гормона роста.
Периодические ритмы, обычно наблюдаемые у морских животных, часто следуют (примерно) 12-часовому переходу от прилива к отливу и обратно.Хрономедицина - это область медицины, в которой используется представление о биологических ритмах, которые изучаются в рамках хронобиологии. Биологические ритмы - это ритмические проявления временной структуры организма, поэтому хрономедицина не исчерпывается одними только биологическими ритмами, а пытается рассмотреть всю «временную структуру организма» в целом.Хрономедицина (как и сама хронобиология) - это молодая область междисциплинарных исследований, которая находится в процессе становления. В хрономедицине находят свое применение методы математической обработки временных рядов, которые используются для анализа ритмических проявлений физиологических процессов организма.Таким образом хрономедицина оказывается на стыке наук: медицины (диагностика и лечение заболеваний), хронобиологии (разработка теоретических представлений) и математики (разработка методов математического анализа ритмических проявлений).Десинхроноз (от латинского de- - приставка, означающая удаление, и греческого synchronos - одновременный) - изменение различных физиологических и психических функций организма в результате нарушения суточных ритмов его функциональных систем. Причины Д.: рассогласование функциональных ритмов организма с показаниями внешних датчиков времени, например, при трансмеридиональных перелётах, перелётах на значительное расстояния в широтном направлении; устойчивое рассогласование по фазе ритма сон - бодрствование (работа в вечерние и ночные смены); частичное или полное отсутствие привычных приборов времени. Признаки Д.: плохой сон, ухудшение аппетита, раздражительность, снижение работоспособности, апатия, вялость. Продолжительность таких расстройств от 1 до 14 дней.
Билет №35. Профилактика наследственных заболеваний и болезней с наследственным предрасположением. Пренатальная диагностика, ее методы и возможности.
Ограниченные возможности лечения наследственных болезней и предсказуемый характер передачи генов от поколения к поколению заставили сосредоточить внимание на профилактике как наиболее надежном и эффективном способе предотвращения этих болезней. Профилактические методы включают генетическое обследование, медико-генетическое консультирование и пренатальную диагностику. Наиболее эффективной мерой профилактики наследственных заболеваний является выявление гетерозиготных носителей мутаций, так как при этом удается предотвратить рождение первого больного ребенка в семьях высокого риска. Родственники больного с большой вероятностью могут быть гетерозиготными носителями мутантных аллелей, поэтому в тех случаях, когда это возможно, они подлежат обследованию в первую очередь. Для болезней, сцепленных с полом, это касается родственников по женской линии - сестер, дочерей и теток пробанда. Их диагностика особенно важна, так как вероятность рождения больных сыновей в потомстве носительниц мутаций очень высока и не зависит от генотипа супруга. При аутосомно-рецессивных заболеваниях половина сибсов родителей и две трети здоровых сибсов больного будут гетерозиготными носителями мутации. Поэтому в тех семьях, где принципиально возможна молекулярная идентификация мутантных аллелей, необходимо обследовать максимальное число родственников больного пробанда для выявления гетерозиготных носителей. Иногда в больших семьях с разветвленными родословными удается проследить наследование неидентифицируемых мутаций с помощью косвенных методов молекулярной диагностики. Для заболеваний, распространенных в определенных популяциях или в каких-то этнических группах и обусловленных присутствием одного или нескольких преобладающих и легко идентифицируемых мутантных аллелей, возможно проведение тотального скрининга на гетерозиготное носительство этих мутаций среди определенных групп населения, например, среди беременных женщин или среди новорожденных. Считается, что подобный скрининг экономически оправдан в том случае, если при проведении процедуры выявляются аллели, составляющие не менее 90-95% всех мутаций данного гена в исследуемой популяции. Выявленные при подобных обследованиях носители мутаций также составляют группу риска, и в последующем должны быть аналогичным образом протестированы их супруги. Однако, даже в том случае, если мутация найдена только у одного из родителей, вероятность рождения больного ребенка несколько выше популяционной частоты, но, конечно, значительно меньше 25%. Конкретное значение этого риска зависит от общей частоты мутаций соответствующего гена в популяции. В таких семьях (по желанию родителей) также может быть проведена пренатальная диагностика и прослежено наследование мутантного аллеля. При отсутствии этой мутации у плода прогноз считается благоприятным, независимо от того, какие аллели ребенок получит от второго супруга.
С профилактической точки зрения всю наследственную патологию целесообразно подразделить на три категории:
1) вновь возникающие мутации (в первую очередь это анеуплоидии и тяжелые формы доминантных мутаций);
2) унаследованные от предыдущих поколений (как генные, так и хромосомные);
3) болезни с наследственной предрасположенностью.
Различают первичную профилактику наследственной патологии и вторичную профилактику наследственной патологии.
Под первичной профилактикой понимают такие меры, которые должны предупредить зачатие или рождение больного ребенка.
Профилактика вновь возникающих мутаций должна сводиться к уменьшению темпа мутационного процесса. Последний же протекает интенсивно.
Современной основой профилактики наследственной патологии являются теоретические разработки в области генетики человека и медицины, которые позволили понять:
1) молекулярную природу наследственных болезней, механизмы и процессы их развития в пре и постнатальном# периоде;
2) закономерности сохранения мутаций (а иногда и распространения) в семьях и популяции;
3) процессы возникновения и становления мутаций в зародышевых и соматических клетках.
Пренатальная диагностика - дородовая диагностика, с целью обнаружения патологии на стадии внутриутробного развития. Позволяет обнаружить более 90 % плодов с синдромом Дауна (трисомия 21); трисомии 18 (известной как синдром Эдвардса) около 97 %, более 40 % нарушений развития сердца и др. В случае наличия у плода болезни родители при помощи врача-консультанта тщательно взвешивают возможности современной медицины и свои собственные в плане реабилитации ребенка. В результате семья принимает решение о судьбе данного ребенка и решает вопрос о продолжении вынашивания или о прерывании беременности. К пренатальной диагностике относится и определение отцовства на ранних сроках беременности, а также определение пола ребенка.
У кузнечика развитие
1) непрямое
2) с куколкой
4) с полным превращением
Пояснение.
У всех насекомых развитие непрямое (с метаморфозом, с превращением). Превращение может быть полное и неполное.
Полное: яйцо, личинка, куколка, взрослое насекомое. Характерно для бабочек (чешуекрылых), жуков (жесткокрылых), комаров и мух (двукрылых), пчел (перепончатокрылых) и т. д.
Неполное: яйцо, личинка, взрослое насекомое (отсутствует стадия куколки). Характерно для кузнечиков и саранчи (прямокрылых).
Итак, у кузнечика развитие непрямое с неполным превращением.
Ответ: 1
Гость
27.05.2012 00:24
У КУЗНЕЧИКА РАЗВИТИЕ ПРЯМОЕ С НЕПОЛНЫМ ПРЕВРАЩЕНИЕМ, ОТВЕТ НОМЕР - 3!!! (Прямое постэмбриональное развитие - это когда родившийся организм отличается от взрослого меньшими размерами и недоразвитием органов. В случае прямого развития молодая особь мало чем отличается от взрослого организма и ведет тот же образ жизни, что и взрослые)
Гость
Гость, Вы ошибаетесь:(
Кузнечик - насекомое - развитие непрямое, без превращения, проходит в 3 стадии.
Постэмбриональное развитие бывает прямым и непрямым.
Прямое развитие - развитие, при котором появившийся организм идентичен по строению взрослому организму, но имеет меньшие размеры и не обладает половой зрелостью. Дальнейшее развитие связано с увеличением размеров и приобретением половой зрелости. Например: развитие рептилий, птиц, млекопитающих.
Непрямое развитие (личиночное развитие, развитие с метаморфозом) - появившийся организм отличается по строению от взрослого организма, обычно устроен проще, может иметь специфические органы, такой зародыш называется личинкой. Личинка питается, растет и со временем личиночные органы заменяются органами, свойственными взрослому организму (имаго). Например: развитие лягушки, некоторых насекомых, различных червей.
Гость
26.10.2012 00:05
Самые древние группы насекомых (тараканы, кузнечики, стрекозы) имеют прямое развитие.
В подтверждение выдержка из энциклопедии:"Кузнечики относятся к насекомым с прямым развитием, это значит, что их личинки внешне похожи на взрослых особей и отличаются только размерами (у других насекомых разница в строении имаго и личинки может быть огромной) и отсутствием крыльев."
http://www.animalsglobe.ru/kuznechiki/
Гость
Аня, можете остаться при своем мнении, но у насекомых - тип развития НЕПРЯМОЕ.
И вот этому сайту я больше доверяю, чем тому, который указываете Вы
http://sbio.info/page.php?id=127
Гость
Да, везде-везде пишут правильно:
непрямое с неполным превращением
Гость
20.02.2013 07:39
У кузнечика неполное развитие. В подтверждение своих слов хочу привести выдержку из "Биологии" Грина ("Мир", 1990, стр. 137-138):
"Как правило, каждая последующая личинка (нимфа или возраст) становится все более похожей на взрослое насекомое. Такой тип развития называется гемиметаболическим метаморфозом. его подразделяют на постепенный метаморфоз, когда нимфы и взрослые формы занимают одни и те же местообитания и питаются одинаковой со взрослыми пищей, и неполный метаморфоз, когда у нимф развиваются адаптивные признаки, позволяющие им осваивать другие местообитания и питаться другой, чем взрослое насекомое пищей."
На стр. 142 приведена таблица, в которой приведены характеристики гемиметаболического метаморфоза:
"Прямое развитие; Наружные зачатки крыльев; Незрелые формы - нимфы, похожие на взрослые формы", а также приведены примеры: отряды Поденки, Тараканы, Настоящие саранчовые.
Рассмотрите рисунки 93 и 94. Какие два типа развития характерны для изображенных на рисунках животных. Какие стадии в своем развитии проходят саранча, бабочка, рыба, лягушка и человек?
Рис. 93. Послезародышевое прямое развитие
Индивидуальное развитие организма продолжается и после его рождения, когда зародыш уже сформировался и может существовать самостоятельно вне яйца или тела матери. Период развития организма после рождения называют после зародышевым., или постэмбриональным (от лат. пост - после и эмбрион). У различных организмов этот период протекает по-разному. Поэтому различают прямое и непрямое развитие.
Прямое и непрямое развитие. Прямое развитие проходит без превращений. Родившийся организм имеет сходство со взрослой особью и отличается только величиной, пропорциями тела и недоразвитием некоторых органов. Такое развитие, в основном, наблюдают у рыб, пресмыкающихся, птиц и млекопитающих (рис. 93). Так, из икринки рыбы выходит личинка с желточным мешком. Она развивается в малька, похожего на взрослую особь, но отличающегося от нее недоразвитием ряда органов.
При развитии с превращением (рис. 94) из яйца появляется личинка совершенно непохожая на взрослый организм. Такое развитие называется непрямым, или развитием с метаморфозом (от греч. метаморфозис - превращение), т. е. с несколькими личиночными стадиями постепенного превращения во взрослую особь. Личинки активно питаются, растут, но, за редким исключением, не способны к размножению.
Рис. 94. Послезаролышевое непрямое развитие (полный метаморфозу бабочки): 1 - яйцо: 2 - личинка (гусеница): 3 - куколка; 4 - взрослое насекомое
Развитие с метаморфозом характерно для насекомых и земноводных. Причем, у насекомых метаморфоз может быть полным и неполным. При развитии с полным метаморфозом насекомые проходят ряд последовательных стадий, как правило, резко отличающихся друг от друга образом жизни и характером питания. Например, у бабочки из яйца выходит гусеница, которая имеет чер-веобразую форму тела. Затем гусеница после нескольких линек превращается в куколку - неподвижную стадию, которая не питается, а только развивается во взрослое насекомое. Через некоторое время из куколки выходит бабочка. Пища и способ питания у личинки и взрослого насекомого различаются. Гусеница ест листья растений и имеет грызущий ротовой аппарат, а бабочка питается нектаром цветков и имеет сосущий ротовой аппарат. Иногда у насекомых некоторых видов взрослая особь вообще не питается, а сразу приступает к размножению (тутовый шелкопряд).
При развитии с неполным метаморфозом стадия куколки отсутствует и личинки мало отличаются от взрослых насекомых. Так, у саранчи вышедшая из яйца личинка имеет по сравнению с взрослой стадией меньшую величину и у нее недоразвиты крылья.
Среди позвоночных животных развитие с превращением наблюдают, в основном, у земноводных. Например, у лягушки личиночная стадия - головастик. По выходу из икринки он напоминает малька рыбы. У него отсутствуют конечности, имеются жабры вместо легких, хвост, при помощи которого он активно плавает в воде. Спустя некоторое время у головастика формируются конечности, развиваются легкие, зарастают жаберные щели и исчезает хвост. Через два месяца после выхода из икринки головастик превращается во взрослую лягушку.
Превращение личинки во взрослую особь связано с выработкой железами внутренней секреции специальных гормонов. Например, для превращения головастика в лягушку необходим гормон щитовидной железы - тироксин. В некоторых случаях при недостатке гормонов личиночный период может затягиваться на всю жизнь и на этой стадии организм может приступить к размножению. Так, личинка земноводного амбистомы - аксолотль при недостатке гормона щитовидной железы не превращается во взрослую особь и может размножаться (рис. 95). При добавлении в воду тироксина развитие идет до конца и аксолотль превращается в амбистому.

Рис. 95. Амбистома (слева) и ее личинка аксолотль (справа)
Рост. Характерное свойство индивидуального развития - рост организма, т. е. увеличение его размеров и массы. По характеру роста всех животных можно разделить на две группы - с неопределенным и определенным ростом. При неопределенном росте размеры тела организма увеличиваются в течение всей его жизни. Это наблюдается, например, у моллюсков, земноводных, рыб и рептилий. Организмы с определенным ростом прекращают расти на определенном этапе развития. Таковы насекомые, птицы и млекопитающие. Темпы роста у животных изменяются в течение всего периода и находятся под контролем гормонов. Например, у млекопитающих (в том числе и у человека) рост регулируется гормоном гипофиза соматотропином. Он активно вырабатывается в детском возрасте, а после полового созревания количество гормона постепенно уменьшается и рост прекращается.
После интенсивного периода роста организм вступает в стадию зрелости, для которой характерно также изменение физиологических процессов в организме. Этот период связан с деторождением.
Старение и смерть. Продолжительность жизни зависит от индивидуальных особенностей вида организма, но не зависит от уровня его организации. Например, мыши живут всего 4 года, ворон - до 70 лет, а моллюск пресноводная жемчужница - до 100 лет.
Процесс индивидуального развития организма заканчивается старением и смертью. Старение - это общебиологическая закономерность, свойственная всем организмам. В процессе старения изменяются все системы органов, нарушаются их структура и функции.
Существует несколько теорий старения. Одна из первых была предложена русским ученым Ильей Ильичей Мечниковым. Согласно этой теории, старение организма связано с усилением в нем процессов интоксикации, самоотравления в результате накопления продуктов обмена веществ и деятельности гнилостных бактерий.
Многие современные теории предполагают, что старение организма - следствие изменений в генетическом аппарате клеток, которые приводят к снижению активности процессов биосинтеза белков. Существенной причиной изменения генетической активности является ослабление работы белков-ферментов. С возрастом повышается частота хромосомных нарушений. Восстановление поврежденных участков ДНК идет медленнее, накапливаются мутации, которые проявляются в структурах РНК и белков.
Высказываются научные гипотезы, которые связывают старение организма с гормональными нарушениями, в частности с изменением функции щитовидной железы.
У человека процессы старения обусловлены действием многих биологических факторов. Немаловажную роль в старении играет и социальная среда, окружающая человека. Наука, занимающаяся проблемами старения человека, называется геронтологией (от греч. герон - старец). Старение - это неизбежный этап развития любого организма. Далее наступает смерть, которая является необходимым условием для продолжения жизни других организмов.
Упражнения по пройденному материалу
- Какие типы после зародышевого развития вам известны?
- В чем разница между прямым и непрямым развитием? Приведите примеры животных с разным типом развития.
- В чем заключается преимущество развития с превращением?
- Чем развитие с полным метаморфозом отличается от развития с неполным метаморфозом? Приведите примеры животных с разным типом метаморфоза.
- В чем заключается старение организма? Какие теории старения вам известны? Какая наиболее вероятна, по вашему мнению? Ответ обоснуйте.
- В чем биологический смысл смерти организма?
В постэмбриональном периоде развития завершается формирование органов тела, происходит половое созревание , размножение , наступает старение и смерть организма.
Все настоящие многоклеточные орга-низмы по типу постэмбрионального раз-вития можно разделить на две группы — с прямым и непрямым типом развития .
Непрямое развитие встречается только у многоклеточных животных. Это те случаи, когда из яйца появляется личин-ка — зародыш, который по своему строе-нию существенно отличается от взрослых особей, но уже способен самостоятельно питаться. Личинка всегда устроена проще и размерами обычно меньше взрослого организма, внешне она похожа на своих далёких предков (например, личинка лягушки — головастик — похожа на небольшую рыб-ку). У неё есть особые личиночные органы, а потому она может вести совершенно иной образ жизни, чем зрелые особи своего вида. У неё нет даже зачаточных половых органов, по которым можно установить, в кого она разовьётся — в самку или самца.
Обязательным условием непрямого развития является наличие метаморфозов (от греч. метаморфосис — превраще-ние) — глубоких преобразований организма, происходящих в течение постэмбрионального периода жизни. У животных этот процесс затрагивает не какие-то отдельные части тела, а организм в целом (рис. 89). В процессе развития личиночные органы исчезают, а их место занимают органы, свойственные взрослому животному.
У растений также могут происходить метаморфозы. Это не только различные видоизменения органов под воздействием окружающей среды, например, отмирание наземной части побе-га в жаркий или холодный период и временное превращение растения в луковицу или корневище, но и метаморфозы органов со сменой их форм и функций. Так, в процессе индивидуального развития у некоторых видов побеги могут превращаться либо в листья, либо в колючки. Характер метаморфоза растений определяют условия окружающей среды. Кроме того, он не затрагивает, как у животных, организм в целом. Именно поэто-му в постэмбриональном развитии растений, даже несмотря на наличие у них метаморфозов, не выделяют личиночных стадий.
Личиночное развитие встречается в большинстве групп животных (губок, членистоногих, червей, моллюсков, кости-стых рыб, земноводных и др.), причём это никак не обусловлено тем, в какой среде — водной или сухопутной — живут особи этих видов во взрослом состоянии.
Почему в своём развитии многие животные обязательно проходят личиночную стадию? Считается, что это связано с де-фицитом питательных веществ, которых не хватает в яйце для завершения зародышевого развития. В результате детёныши появляются на свет недоразвитыми, по сути, будучи ещё заро-дышами. Такое объяснение подтверждается сравнением размеров яиц позвоночных с личиночными стадиями разви-тия (костистые рыбы и земноводные) с яйцами рептилий и птиц, у которых такие стадии отсутствуют. У самок рептилий и птиц яйца огромны и богаты желтком, которого хватает для полноценного развития зародыша. У рыб же и земновод-ных яйца (икра) небольшие, запасательных веществ в них не очень много. Гигантские яйцеклетки свойственны ещё одним безличиночным животным — хрящевым рыбам. Личинки не встречаются и у живородящих животных (некоторых акул, морских змей, млекопитающих и ряда других). Очевидно, у этих видов материнский организм обладает достаточными ресурсами для того, чтобы их по-томство завершило зародышевое развитие внутриутробно.
Важным преимуществом личиночного развития счита-ют то обстоятельство, что зачастую личинки и взрослые особи обитают в разных средах (в воде проис-ходит развитие крылатых насекомых: стрекоз, подёнок, комаров, слепней и многих других). Считается, что таким образом устраняется конкуренция между собой особей одного вида за пищевые ресурсы. Материал с сайта
Личинки бывают настолько не похожи на взрослые формы, что вначале их часто описывали как представителей разных родов, отрядов, классов и даже типов царства животных (рис. 90). С тех пор многие личинки сохранили свои собственные названия: наяда в древнегреческой мифологии — нимфа лесных ручьёв — личинка стрекозы, пескоройка — личинка миноги, живущая в норах, вырытых в иле; трохофоры (от греч. трохос — колесо и форос — несущий) — свободно плавающие личинки некоторых групп моллюсков и кольчатых червей; науплиусы (от греч. науплиус — животное с панцирем) — ранние личинки многих ракообразных; лептоцефал (по лат. короткоголов) — личинка угря, для которой характерно совершенно прозрачное тело и другие.
После появления организма на свет начинается его постэмбриональное развитие (постнатальное для человека), которое у разных организмов протекает от нескольких дней до сотен лет в зависимости от их видовой принадлежности. Следовательно, продолжительность жизни - это видовой признак организмов, не зависящий от уровня их организации.
В постэмбриональном онтогенезе различают ювениальный и пубертатный периоды, а также период старости, заканчивающийся смертью.
Ювенильный период. Этот период (от лат. juvenilis - юный) определяется временем от рождения организма до полового созревания. У разных организмов он протекает по-разному и зависит от типа онтогенеза организмов. Для этого периода характерно либо прямое, либо непрямое развитие.
В случае организмов, для которых характерно прямое развитие (многие беспозвоночные, рыбы, пресмыкающиеся, птицы, млекопитающие, человек), вылупившиеся из яйцевых оболочек или новорожденные сходны со взрослыми формами, отличаясь от последних лишь меньшими размерами, а также недоразвитием отдельных органов и несовершенными пропорциями тела.
Характерной особенностью роста в ювенильный период организмов, подверженных прямому развитию, является то, что происходит увеличение количества и размеров клеток, изменяются пропорции тела. Рост человека в разные периоды его онтогенеза. Рост разных органов человека не-равномерен. Например, рост головы заканчивается в детстве, ноги достигают пропорциональной величины примерно к 10 годам. Наружные половые органы очень быстро растут в возрасте 12-14 лет. Различают определенный и неопределенный рост. Определенный рост характерен для организмов, которые к определенному возрасту прекращают свой рост, например, насекомые, млекопитающие, человек. Неопределенный рост характерен для организмов, которые растут всю жизнь, например, моллюски, рыбы, земноводные, рептилии, многие виды растений.
В случае непрямого развития организмы претерпевают превращения, называемые метаморфозами (от лат. metamorphosis - превращение). Они представляют собой видоизменения организмов в процессе развития. Метаморфозы широко встречаются у кишечнополостных (гидры, медузы, коралловые полипы), плоских червей (фасциолы), круглых червей (аскариды), моллюсков (устрицы, мидии, осьминоги), членистоногих (раки, речные крабы, омары, кре-ветки, скорпионы, пауки, клещи, насекомые) и даже у некоторых хордовых (оболочечники и земноводные). При этом различают полные и неполные метаморфозы. Наиболее выразительные формы метаморфозов наблюдают у насекомых, которые подвергаются как неполным, так и полным метаморфозам.
Неполное превращение - это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитипизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов.
Полное превращение - это такое развитие, при котором из яйцевых оболочек освобождается личинка, существенно отличающаяся по строению от взрослых особей. Например, у бабочек и многих насекомых личинками являются гусеницы. Гусеницы подвержены линьке, причем могут линять по не-скольку раз, превращаясь затем в куколки. Из последних развиваются взрослые формы (имаго), которые не отличаются от исходных.
У позвоночных метаморфозы встречаются среди земноводных и костных рыб. Для личиночной стадии характерно наличие провизорных органов, которые либо повторяют признаки предков, либо имеют явно приспособительное значение. Например, для головастика, являющегося личиночной формой лягушки и повторяющего признаки исходной формы, характерны рыбообразная форма, наличие жаберного дыхания, одного круга кровообращения. Приспособительными признаками головастиков являются их присоски, длинный кишечник. Для личиночных форм характерно также и то, что по сравнению со взрослыми формами, они оказываются приспособленными к жизни в совершенно иных условиях, занимая другую экологическую нишу и другое место в цепи питания. Например, личинки лягушек имеют жаберное дыхание, тогда как взрослые формы - легочное. В отличие от взрослых форм, которые являются плотоядными существами, личинки лягушек питаются растительной пищей.
Последовательность событий в развитии организмов часто называют жизненными циклами, которые могут быть простыми и сложными. Наиболее простые циклы развития характерны, например, для млекопитающих, когда из оплодотворенной яйцеклетки развивается организм, который снова продуцирует яйцеклетки и т. д. Сложными биологическими циклами являются циклы у животных, для которых характерно развитие с метаморфозами.
Развитие и дифференциация, связанные с метаморфозами, являются результатом естественного отбора, благодаря которому многие личиночные формы, например, гусеницы насекомых и головастики лягушек адаптированы к среде лучше, чем взрослые половозрелые формы.
Пубертатный период. Этот период называют еще зрелым, и он связан с половой зрелостью организмов. Развитие организмов в этот период достигает максимума.
На рост и развитие в постэмбриональный период большое влияние оказывают факторы среды. Для растений решающими факторами являются свет, влажность, температура, количество и качество питательных веществ в почве. Для животных первостепенное значение имеет полноценное кормление (наличие в корме белков, углеводов, липидов, минеральных солей, витаминов, микроэлементов). Важны также кислород, температура, свет (синтез витамина Д).
Рост и индивидуальное развитие животных организмов подвержены нейрогуморальной регуляции со стороны гуморальных и нервных механизмов регуляции. У растений обнаружены гормон - подобные активные вещества, получившие название фитогормонов. Последние влияют на жизненно важные отправления растительных организмов.
В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных вырабатывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности, те, которые имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируется гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлена гормональная регуляция созревания яиц, протекание диапаузы.
У позвоночных железами внутренней секреции являются гипофиз, эпифиз, щитовидная, паращитовидная, поджелудочная, надпочечники и половые железы, которые тесно связаны одна с другой. Гипофиз у позвоночных вырабатывает гонадотропный гормон, стимулирующий деятельность половых желез. У человека гормон гипофиза влияет на рост. При недостатке развивается карликовость, при избытке - гигантизм. Эпифиз продуцирует гормон, который влияет на сезонные колебания в половой активности животных. Гормон щитовидной железы влияет на метаморфоз насекомых и земноводных. У млекопитающих недоразвитие щитовидной железы ведет к задержке роста, недоразвитию половых органов. У человека из-за дефекта щитовидной железы задерживается окостенение, рост (карликовость), не наступает полового созревания, останавливается психическое развитие (кретинизм). Надпочечники продуцируют гормоны, оказывающие влияние на метаболизм, рост и дифференцировку клеток. Половые железы продуцируют половые гормоны, которые определяют вторичные половые признаки. Удаление половых желез ведет к необратимым изменениям ряда признаков. Например, у кастрированных петухов прекращается рост гребня, теряется половой инстинкт. Кастрированный мужчина преобретает внешнее сходство с женщиной (не растет борода и волосы на коже, отлагается жир на груди и в области таза, сохраняется тембр голоса и т. д.).
Фитогормонами растений являются ауксины, цитокинины и гиббереллины. Они регулируют рост и деление клеток, образование новых корней, развитие цветков и другие свойства у растений.
На всех периодах онтогенеза организмы способны к восстановлению утраченных или поврежденных частей тела. Это свойство организмов носит название регенерации, которая бывает физиологической и репаративной.
Физиологическая регенерация - это замена утерянных частей тела в процессе жизнедеятельности организма. Регенерации этого типа очень распространены в животном мире. Например, у членистоногих она представлена линькой, которая связана с ростом. У рептилий регенерация выражается в замещении хвоста и чешуи, у птиц - перьев, когтей и шпор. У млекопитающих примером физиологической регенерации может быть ежегодное сбрасывание оленями рогов.
Репаративная регенерация - это восстановление части тела организма, отторгнутой насильственным путем. Регенерация этого типа возможна у многих животных, но ее проявления различны. Например, она часта у гидр и связана с размножением последних, поскольку из части регенерирует весь организм. У других организмов регенерации проявляются в виде способности отдельных органов к восстановлению после утраты ими какой-либо части. У человека достаточно высокой регенеративной способностью обладают эпителиальная, соединительная, мышечная и костная ткани.
Растения многих видов также способны к регенерации. Данные о регенерации имеют большое значение не только в биологии. Их широко используют в сельском хозяйстве, в медицине, в частности, в хирургии.
Старость как этап онтогенеза. Старость является предпоследним этапом онтогенеза животных, причем ее длительность определяется общей продолжительностью жизни, которая является видовым признаком и которая у разных животных является разной. Наиболее точно старость изучена у человека.
Известны самые различные определения старости человека. В частности, одно из наиболее популярных определений заключается в том, что старость есть накопление последовательных изменений, сопровождающих повышение возраста организма и увеличивающих вероятность его болезней или смерти. Науку о старости человека называют геронтологией (от греч. geron - старик, старец, logos - наука). Ее задачей является изучение закономерностей возрастного перехода между зрелостью и смертью.
Хордовые - один из крупнейших типов животного царства, представители которого освоили все среды обитания. В состав этого типа входят три группы (подтипа) организмов: оболочники (в том числе живущие на дне морские сидячие организмы - асцидии), бесчерепные (похожие на маленьких рыбок морские существа - ланцетники), позвоночные (хрящевые и костистые рыбы, земноводные, пресмыкающиеся, птицы и млекопитающие). Человек - тоже представитель типа хордовых. Происхождение типа хордовых - это важнейший этап в историческом развитии животного мира, означающий появление группы животных с уникальным планом строения, который позволил в дальнейшей эволюции достичь максимальной среди живых существ сложности строения и поведения. В сущности это означает реконструкцию первых шагов на том пути, который привел к человеку. Вот почему проблема происхождения хордовых уже более полутора столетий вызывает большой интерес биологов.
Происхождение хордовых относится к тому периоду эволюции животного царства, от которого сохранилось очень мало палеонтологических остатков. Известно лишь, что в среднем кембрии (около 550 млн. лет назад) уже существовали бесчерепные, родственные современным ланцетникам. Более ранние этапы эволюции хордовых приходится реконструировать на основе изучения современных организмов, преимущественно сравнительно-анатомическими и эмбриологическими методами, привлекая данные и из других областей биологии.
ОСНОВНЫЕ ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ХОРДОВЫХ
Одна из первых концепций, связывающих план строения хордовых с планами строения других групп беспозвоночных, была разработана выдающимся французским сравнительным анатомом Жоффруа Сент-Илером еще в первой половине XIX века. Он считал, что членистые животные (кольчатые черви и членистоногие) могут рассматриваться как перевернутые позвоночные. "Взгляните на рака, опрокинутого на спину, говорил он, - и вы увидите, что различные системы его расположены совершенно так же, как у высших позвоночных животных". Именно эта концепция была в центре публичного спора между Жоффруа Сент-Илером и Жоржем Кювье, который состоялся в Париже в 1830 году в дни июльской революции, свергнувшей Бурбонов. Как известно, в том знаменитом споре победил Жорж Кювье. Однако к идее сравнения хордовых (и позвоночных) и перевернутых кольчатых червей или членистоногих зоологи возвращались неоднократно на протяжении всего XIX века.
Эту точку зрения отстаивал выдающийся зоолог и, как бы мы сейчас сказали, "организатор науки" Антон Дорн, построивший на свои личные средства первую в мире и до сих пор успешно работающую Неаполитанскую зоологическую станцию. В начале XX века идею происхождения хордовых от перевернувшихся на спинную сторону членистоногих отстаивали Гаскелл, Паттен и другие биологи.
Остроумные и хорошо объясняющие план строения хордовых гипотезы их происхождения от перевернутых аннелид или членистоногих утратили свою популярность прежде всего в связи с тем, что стали ясны фундаментальные эмбриологические отличия между кольчатыми червями и членистоногими, с одной стороны, и хордовыми - с другой.
В начале нынешнего столетия были разработаны гипотезы происхождения хордовых от кишечнодышащих - обитающих в толще грунт морских червеобразных организмов. У этих животных (как и у хордовых) имеются жаберные щели, а в передней части тела - опорный вырост кишечника (стомохорд), который сравнивали с хордой. Однако кишечнодышащие резко отличались от хордовых обратным направления тока крови, расположением сердца, строением нервной системы и другими важными особенностями организации. Делались попытки выводить хордовых от иглокожих, немертин, моллюсков и других групп беспозвоночных, однако ни одна из них не имела успеха.
Своеобразной реакцией на неудачи зоологов решить проблему происхождения хордовых традиционными способами явилась разработка так называемых неотенических гипотез, выводящих хордовых от личинок асцидий. Взрослые асцидии - прикрепленные морские животные, фильтрующие воду через объемистый жаберный мешок и улавливающие взвешенные в воде мельчайшие органические частицы. Они не имеют ни хорды, ни нервной трубки и совсем не похожи на хордовых животных. Но, как давно известно (а впервые это показал еще в конце прошлого века выдающийся русский биолог А.О. Ковалевский), личинки асцидий имеют и хорду, и нервную трубку, которые утрачиваются при метаморфозе. На основании этого многие биологи (Берилл, Уитер, Боун и др.) предполагают, что хордовые произошли от ставших половозрелыми личинок асцидий. Правда, в этом случае остается непонятным, откуда взялись сами асцидии. Да и трудно представить, чтобы хорда и нервная трубка возникли у живущих 1 - 2 дня и не питающихся личинок. Более вероятно, что эти особенности - наследие свободноживущих предков асцидий, об организации которых неотенические гипотезы ничего не говорят. Ни одна из рассмотренных гипотез в настоящее время не может считаться истинной, но в каждой из них содержатся отдельные элементы, которые могут оказаться полезными при реконструкции путей происхождения хордовых.
Жизненный цикл клетки: просинтетическая, синтетическая и постсинтетическая фазы. Значение этих фаз в жизни клетки.
Жизн.цикл кл. – это время существования клетки от деления от деления или до смерти. У одноклеточного клеточного цикла совпадает с жизнью особи. В непрерывно размножающих тканевых клетках жизненный цикл клетки совпадает с жизнью особи. В непрерывно размножающихся тканевых клетках жизненный клетки совпадает с митотическим циклом (митоз). Состоит из 3х периодов: пресинтетический период (ПП) –после завершения митоза клетка может вступить в период подготовки к синтезу ДНК. ПП обозначаемый символом G1 (интервал). В этот период в клетке образ-ся РНК и белки, повышается активность ферментов, кот-е участвуют в биосинтезе ДНК, происходит удвоение органоидов и рост клетки. После завершения фазы клетка приступает к синтезу ДНК, т.е. ее репликации – удвоению (в S фазе). Синтетический период (СП) – процесс репликации начинается одновременно во многих точках каждой из хромосом. 2 цепи исходной молекулы ДНК расходятся, и каждая становится матрицей для новых цепей. Каждая дочерняя молекула включает одну старую и одну новую цепь последовательно соединенных нуклеотидов. Продолжительность S фазы в разных клетках неодинакова и составляет 6-12 ч. (в клетках млекопит.). В результате удвоения ДНК в каждой из хромосом оказывается в 2 больше ДНК чем было до начала S фазы, но число хромосом не изменяется, следовательно, после завершения редупликации ДНК хромосомный набор клетки станов-ся 2n4c. Постсинтетический период – после завершения синтеза ДНК – S фазы – клетка не сразу начинает делиться. Время от окончания синтеза ДНК и до начала митоза – это постсинтетический период – G2. В этот период завершается подготовка клетки к митозу, удваиваются белки, завершается рост клетки, накапливается АТФ.
Сцепленное наследования и кроссинговер. Генетические карты, принцип их построения у эукариот.
Сцепленное наследие. Кроссинговер. Генетические карты. Гены кот. входят в состав одной хромосомы называются сцепленными и образуют гр. сцепления. Наследуются они как единое целое т.к. это определяется поведением хромосомы в мейозе. В таком случае расщепление по сцепленным признакам не подчиняется закону независимого наследования. Если гены расположены близко др. к др., то они всегда сохраняются в исходных сочетаниях. Более распространенными явл. ситуации, когда гены располагаются на некот. удалении др. от др. В таком случае частичного сцепления они могут разделяться в результате процесса – кросссинговера – это вид генет-кой рекомбинации. Кроссинговер осуществляется в профазе первого мейотического деления в момент коньюгации хромосом (в это время хроматиды гомологичных хромосом обмениваются фрагментами наследственного материала и появляются новые комбинации генов). Величина кроссинговера, вычисляемая как % отношения рекомбинантов к общей численности потомства, явл. показателем расстояния м/у генами и используется для картирования хромосом – расположения генов на карте хромосомы в строго определенном порядке и на фиксированных расстояниях.
Генетика. Мутации как основы возникновения наследственной изменчивости организмов. Виды мутаций (генные, хромосомные и геномные). Наследственная изменчивость- процесс возникновения наследственных изменений организма - мутации. (в 1899 г. рус. ученым С. И. Коржинским и в 1900-1901 гг. гол. генетиком Г. де Фризом, к-рьш, в частности, ввел термины <мутация» (лат. изменение) и «мутагенез». Способность мутировать присуща всем формам жизни на Земле и лежит в основе фундаментального свойства живого - изменчивости. Сущность мутаций состоит в изменении структуры ДНК, являющейся носителем генетической информации. Мутации приводят к возникновению нового признака или изменению (утрате) старого. Различают спонтанный (возникающий в естественных условиях без четко регистрируемых внешних воздействий) и индуцированный (в результате искусственных воздействий) М.
Виды мутаций: Геномные : - полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3n, 4n, 6n и т. д.) наборами хромосом) и анеуплоидия (гетероплоидия) - изменение числа хромосом, не кратное гаплоидному набору (см. Инге-Вечтомов, 1989). В зависимости от происхождения хромосомных наборов среди полиплоидов различают аллополиплоидов, у которых имеются наборы хромосом, полученные при гибридизации от разных видов, и аутополиплоидов, у которых происходит увеличение числа наборов хромосом собственного генома, кратное n.
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай - объединение целых хромосом, т. н. Робертсоновская транслокация, которая является переходным вариантом от хромосомной мутации к геномной).
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях.
Зоология позвоночных. Происхождение и эволюция класса рыбы. наиболее древние известные бесчелюстные животные, напоминающие рыб, известны из раннего ордовика (около 450-470 миллионов лет назад). С развитием челюстей из одной из жаберных дуг возникли первые рыбы. Кроме челюстей, рыбы имеют парные плавники, внутреннее ухо с тремя полукружными каналами и жаберные дуги. Несмотря на появление первых челюстноротых рыб ещё в ордовике, они занимали подчиненное положение вплоть до девона. Таким образом, рыбы и бесчелюстныесуществовали больше 100 миллионов лет в условиях преобладания бесчелюстных, в отличие от настоящего времени. Хрящевые рыбы появились на рубеже силура и девона, около 420 миллионов лет назад, и достигли расцвета в карбоне. Хрящевые рыбы. Впервые появились панцирные рыбы. 450 млн.лет назад. Первые хордовые животные у которых жаберные аппараты их было 10 или более возле головы – подвижно соединены между собой (до самого конца головы). Из 3-4 пары челюстной аппарат (хватательные). На коже появляются флаксидная чешуя появляется на 3,4 жаберных дугах-удержание пищи, 1-2 дуг –хватательные и удерживающие – так появились подъязычный аппарат от других дуг.
Первые рыбы – акантодии-околодонные,из них произошли костные и хрящевые.Хрящевые (древн.) происхождение в пресных водоемах. Скелет хрящевый, имеется хорда,членистый скелет. Костные рыбы обитают в мировых океанах как минимум с девона; не исключено, что они существовали уже в силуре. Guiyu oneiros - самая ранняя известная костная рыбаКостные рыбы – обладают костным скелетом. Окостенение, образование костной ткани, членистого столба. Строение костного скелета прогрессивный. Костные рыбы обитают в любой среде (пресной,морской,пещерные).
Цитология и гистология. Мембранные системы клетки и мембранный принцип ее организации. Структура и свойства биологических мембран и их свойства. Органоиды клетки: 1)-обязательные-обеспечивают жизнедеятельность клетки -->митохондрии рибосомы плазмалемма аппарат гольджи генетический аппарат.
-->мембранные(одномембранные-аппарат гольджи, вакуоль,гранулярная ЭПР,лизосомы,микросомы) - (двумембранные-митохондрии, пластиды,ядро)
à немембранные (рибосомы,клеточный центр,микротрубочки,микрофиламенты,микрофибриллы)
2) факультативные-узкоспециализированные необязательны для всех клеток àхлоропласты,вакуоль,клеточный центр
Эндоплазматическая сеть-это разветвленная сеть образованная мембраной (одномембранная) на котором нах-ся ферменты.
ЭПС:гладкая (агрокулярные), шероховатые (гранулярные)
На мембранах шероховатая ЭПС нах-ся рибосомы. Функции синтез белков.
Аппарат гольджи-скопление мембранных структур в виде уплощенных цистерн и мелких пузырьков. Цистерны соединяются с каналами эпс. Синтезируются на мембранах эпс белки, полисахариды и жиры транспортируются в А.Г. конденсируются внутри его структур и упаковываются в виде секрета или используются самой клеткой.
Лизосомы – одномембранные органоиды расщепляют белки, полисахаридов,липидов итд. Функция переваривания веществ.
Ядро – двумембранный органоид. Состоит из ядерной мембраны хроматина (ядрышка) кариоплазмы (ядерный сок). На наружной мембране большое количество лизосом. Ядерная мембрана переходит в эпс шероховатую. Оболочка ядра пронизана порами. Поры образуются за счет слияния двух мембран. В состав хроматина входит ДНК в комплексе с белком. Хромосомы образуются только в делящихся клетках за счет спирализации хроматиновых нитей. Функции ядра: хранение информации и ее реализация, передача.
Биологическая мембрана – это тонкая липопротеидная полупроницаемая пленка, состоит из двойного слоя липидных молекул, в которой включены молекулы белка.
Жидкостно-мозаичная модель биологических мембран. Мембрана образует двойной слой фосфолипидов. Фосфолипиды состоят: - головка-полярны (-) нейроны,гидрофильные; хвосты-неполярные,незаряженные,гидрофильные.
Молекулы белка не образуют,располагаются в слое липидов. Большинство белков не связаны с липидами.
Типы белков: -интегральны-пронизывают толщу мембраны; -полуинтегральные-белки наполовину погружаются в мембрану; - периферийные-белки находятся на поверхности мембран.
На долю липидов приходится 60%, белки 40-75%. В состав многих мембран входят углеводы которые образуют слой гликкокаликс имеются только у животных клеток.
Свойства клеточной (биологической)мембраны.
1) Замкнутость – в процессе сборки липидов замки замыкаются сами по себе. Это приводит к образованию закрытых отсеков в клетке которые называются компартменты
2) Динамичность – некоторые молекулы липидов и белков могут перемещаться в мембране т.е. сохраняют способность в диффузии
3) Ассиметричность-различаются наличием гликокаликса, по составу липидов,белков.
4) Избирательная проницаемость – это свойство обеспечивает регуляцию проникновения в клетку необходимых молекул,а также удаление продуктов метаболизма
Функции клеточной мембраны
Барьерная –отграничивает содержимое клетки от внешней среды,т.е. поддержание целостности
Формирование мембранных структур клетки
Транспорт питательных веществ, внутрь отходов наружу
Рецепторная – восприятие сигналов и передача их внутрь клетки
Принимают участие в построении специальных отростков клетки, таких как микроворсинки, реснички, рецепторные выросты.


