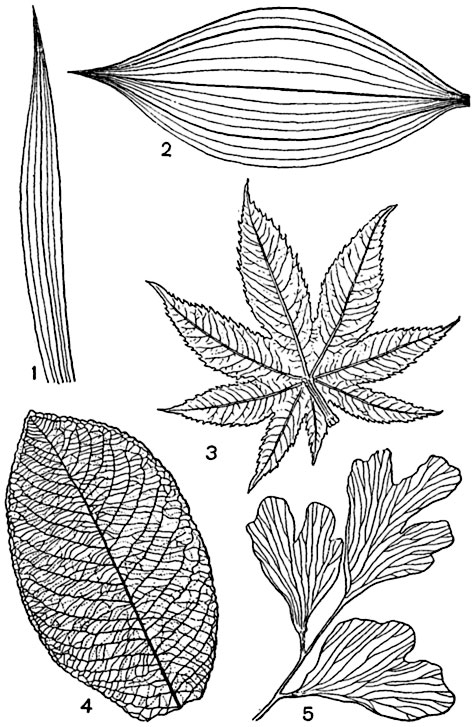
Как устроены устьица. Покровные ткани
ТКАНИ. КЛАССИФИКАЦИЯ ТКАНЕЙ.
В основе организации высших растений лежит принцип специализации клеток, который заключается в том, что каждая клетка организма выполняет не все присущие ей функции, а только некоторые, но зато более полно и совершенно.
Ткани - устойчивые, закономерно повторяющиеся комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций.
Существуют различные классификации тканей, но все они достаточно условны.
В зависимости от основной функции различают несколько групп растительных тканей.
1. Образовательные ткани, илимеристемы, - обладают способностью к делению и формированию всех прочих тканей.
2. Покровные ткани:
Первичные;
Вторичные;
Третичные.
3. Основные ткани - составляют большую часть тела растения. Различают следующие основные ткани:
Ассимиляционные (хлорофиллоносные);
Запасающие;
Воздухоносные (аэренхима);
Водоносные.
4. Механические ткани (опорные, скелетные):
Колленхима;
Склеренхима.
5. Проводящие ткани:
Ксилема (древесина) - ткань восходящего тока;
Флоэма (луб) - ткань нисходящего тока.
6. Выделительные ткани:
Наружные:
Железистые волоски;
Гидатоды - водяные устьица;
Нектарники;
Внутренние:
Выделительные клетки с эфирными маслами, смолами, дубильными веществами;
Многоклеточные вместилища выделений, млечники.
По способности клеток к делению различают два типа тканей: образовательные, илимеристемы, ипостоянные - покровные, выделительные, основные, механические, проводящие.
Ткань называют простой, если все ее клетки одинаковы по форме и функциям (паренхима, склеренхима, колленхима).Сложные ткани состоят из клеток, неодинаковых по форме, строению и функциям, но связанных общим происхождением (например, ксилема, флоэма).
Существует также классификация тканей, основанная на их происхождении (онтогенетическая). Согласно этой классификации различают первичные и вторичные ткани. Из первичной меристемы, находящейся на верхушке побега и кончике корня, а также из зародыша семени формируются первичные постоянные ткани (эпидерма, колленхима, склеренхима, ассимиляционная ткань, эпиблема). Клетки постоянных тканей неспособны к дальнейшему делению. Из клеток специализированной меристемы -прокамбия - формируютсяпервичные проводящие ткани (первичная ксилема, первичная флоэма).
Из вторичной меристемы - камбия - формируютсявторичные ткани: вторичная ксилема, вторичная флоэма; изфеллогена образуются пробка, феллодерма, чечевички, возникающие при утолщении стебля и корня. Вторичные ткани, как правило, встречаются у голосеменных и двудольных покрытосеменных растений. Мощное развитие вторичных тканей - древесины и луба характерно для древесных растений.
ОБРАЗОВАТЕЛЬНЫЕ ТКАНИ
Образовательные ткани благодаря постоянному митотическому делению их клеток обеспечивают образование всех тканей растения, т.е. фактически формируют его тело. Любая клетка в своем развитии проходит три стадии: эмбриональную, роста и стадию дифференциации (то есть приобретения клеткой определенной функции). По мере дифференциации зародыша первичная меристема сохраняется только на верхушке будущего побега (в конусе нарастания) и на кончике корня - апикальные (верхушечные)меристемы. Зародыш любого растения состоит из клеток меристемы.
Цитологическая характеристика меристем. Типичные признаки наиболее отчетливо выражены в верхушечных меристемах. Эти меристемы составлены изодиаметрическими многогранными клетками, не разделенными межклетниками. Их оболочки тонки, содержат мало целлюлозы и способны растягиваться.
Полость каждой клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение, и интенсивно делящейся митозом. В гиалоплазме много диффузно разбросанных рибосом, пропластид, митохондрий и диктиосом. Вакуолей немного, и они мелкие. Проводящие ткани образуются из меристемы, имеющей прозенхимную форму и крупные вакуоли, - прокамбия и камбия. Клетки прокамбия в поперечном сечении многоугольные, клетки камбия - прямоугольные.
Клетки, сохраняющие свои меристематические свойства, продолжают делиться, образуя все новые и новые клетки, называемые инициалями. Часть дочерних клеток дифференцируется, превращаясь в клетки различных тканей, их называютпроизводными инициалей. Клетки инициалей могут делиться неопределенно много раз, а производные инициалей делятся один или несколько раз и превращаются в постоянные ткани.
По происхождению различают первичные и вторичные меристемы.
Первичные меристемы
Первичные меристемы происходят непосредственно из меристемы зародыша и обладают способностью к делению. По положению в растении первичные меристемы могут быть верхушечными (апикальными), вставочными (интеркалярными) и боковыми (латеральными).
Верхушечные (апикальные) меристемы - такие меристемы, которые располагаются у взрослых растений на верхушках стеблей и кончиках корней и обеспечивают рост тела в длину. У стеблей в конусе нарастания выделяют два меристематических слоя: тунику, из которой образуются покровная ткань и периферическая часть первичной коры, и корпус, из которого образуются внутренняя часть первичной коры и центральный осевой цилиндр (рис. 2.3).
Рис. 2.3. Апикальные меристемы стебля:а - продольный срез: 1 - конус нарастания; 2 - зачаток листа; 3 - бугорок пазушной почки;
В кончике корня различают три слоя:
1) дерматоген, из которого образуется первичная покровно-всасывающая ткань - ризодерма;
2) периблему, из которой развиваются ткани первичной коры;
3) плером, образующий ткани центрального осевого цилиндра.
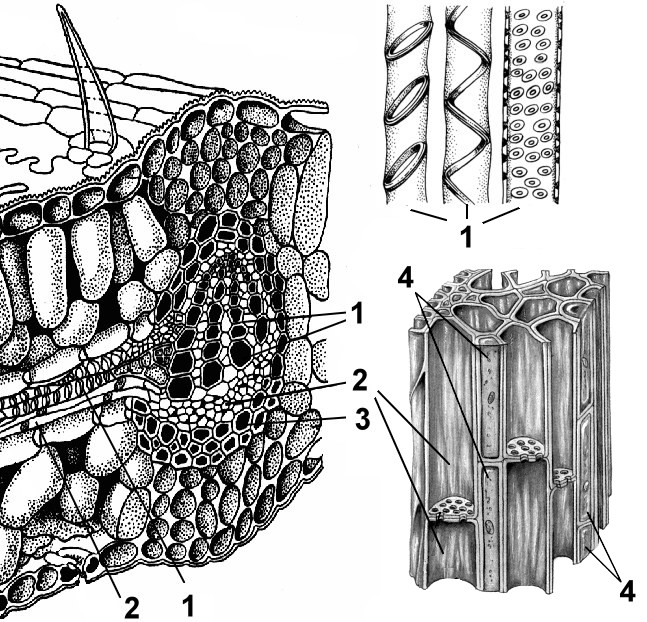
Боковые (латеральные) меристемы по происхождению могут быть первичными и вторичными, на поперечном срезе осевых органов имеют вид колец. Примером первичной боковой меристемы служат прокамбий и перицикл. Изпрокамбия формируются камбий и первичные элементы сосудисто-волокнистых пучков (первичная флоэма и первичная ксилема), при этом клетки прокамбия непосредственно дифференцируются в клетки первичных проводящих тканей.
Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Вставочные (интеркалярные) меристемы чаще первичные и сохраняются в виде отдельных участков в зонах активного роста в различных частях растения (например, в основании черешков листьев, у оснований междоузлий). В основании междоузлий у злаков деятельность этой меристемы ведет к удлинению междоузлий, что обеспечивает рост стебля в длину.
Вторичные меристемы
К вторичным относят боковые и раневые меристемы.
Боковые (латеральные) меристемы представленыкамбием ифеллогеном. Они формируются из промеристем (прокамбия) или постоянных тканей путем их дедифференцировки. Клетки камбия делятся перегородками, параллельными поверхности органа (периклинально). Из клеток, отложенных камбием наружу, развиваются элементы вторичной флоэмы, а из клеток, отложенных внутрь, - вторичной ксилемы. Камбий, возникший из постоянных тканей путем дедифференцировки, называютдобавочньм. По строению и функции он не отличается от камбия, возникшего из промеристем. Феллоген формируется из постоянных тканей, расположенных в субэпидермальных слоях (под эпидермой). Делясь периклинально, феллоген отделяет наружу будущие клетки пробки (феллемы), а внутрь - клетки феллодермы. Таким образом, феллоген формирует вторичную покровную ткань - перидерму. Боковые меристемы располагаются параллельно поверхности органа и обеспечивают рост осевых органов в толщину.
Раневые меристемы образуются при повреждении тканей и органов. Вокруг повреждения живые клетки дедифференцируются, начинают делиться и тем самым превращаются во вторичную меристему. Их задача - образовать плотную защитную ткань, состоящую из паренхимных клеток, -каллюс. Эта ткань беловатого или желтоватого цвета, ее клетки имеют крупные ядра и достаточно толстые клеточные стенки. Каллюс возникает при прививках, обеспечивая срастание привоя с подвоем, и в основании черенков. В нем могут закладываться придаточные корни и почки, поэтому его используют для получения культуры изолированных тканей.
ПОКРОВНЫЕ ТКАНИ
Первичная покровная ткань
К первичной покровной ткани относят эпидерму собственно эпидермальных, околоустьичных, замыкающих клеток устьица и трихом.
Пектиновые вещества и целлюлоза, входящие в клеточную стенку, могут подвергаться ослизнению с образованием слизей икамедей. Они представляют собой полимерные углеводы, родственные пектиновым веществам, и отличаются способностью к сильному набуханию при соприкосновении с водой. Камеди в набухшем состоянии клейкие и могут вытягиваться в нити, тогда как слизи сильно расплываются и в нити не вытягиваются. Пектиновые слизи встречаются у представителей семейств лилейных, крестоцветных, мальвовых, липовых, розоцветных, в отличие от целлюлозных слизей, встречающихся гораздо реже (например, у орхидных).
Устьица
представляют собой
высокоспециализированные образования
эпидермы, состоящие из двух замыкающих
клеток бобовидной формы и устьичной
щели (своеобразного межклетника между
ними). Имеются главным образом в листьях,
но встречаются и в стебле (рис. 2.6).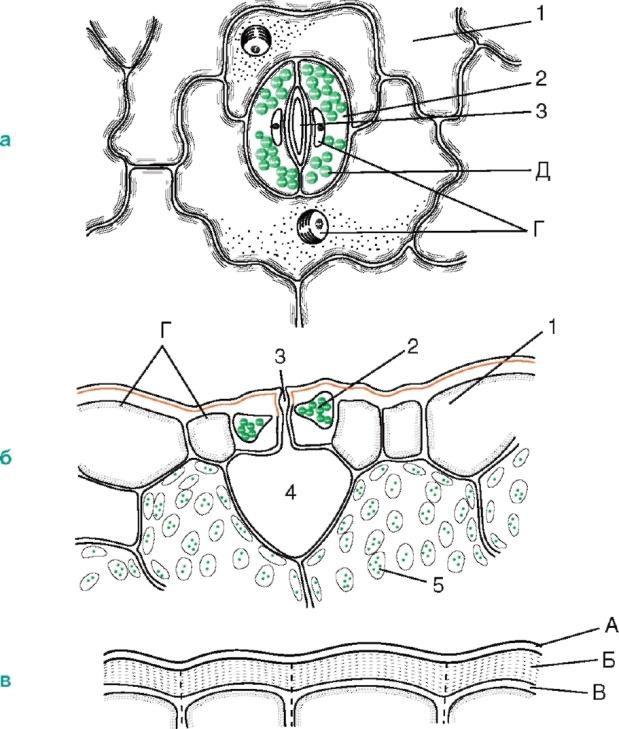
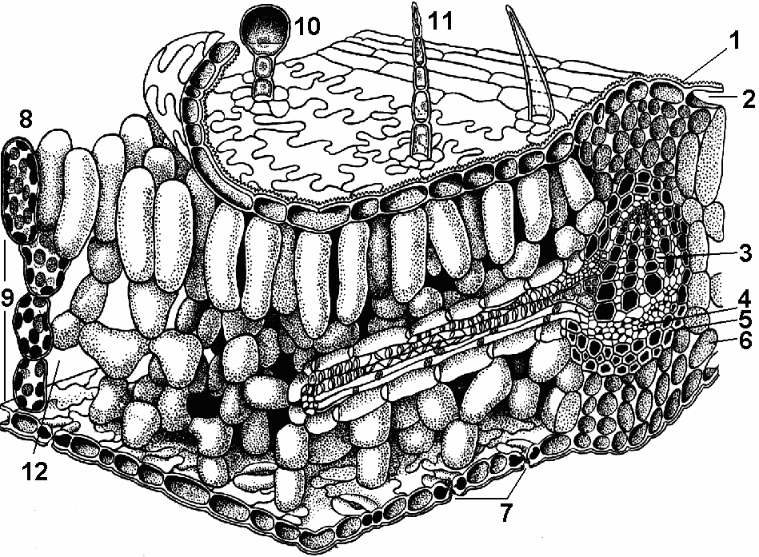
Рис. 2.6. Строение устьица:а, б - кожица листа тимьяна (вид сверху и на поперечном срезе);в - кожица со стебля цереуса (семейство кактусовых); 1 - собственно эпидермальные клетки; 2 - замыкающие клетки устьица; 3 - устьичная щель; 4 - воздухоносная полость; 5 - клетки хлорофиллоносной паренхимы; А - кутикула; Б - кутикулярный слой - оболочка с суберином и воском; В - целлюлозный слой стенки; Г - ядро с ядрышком; Д - хлоропласты
Стенки замыкающих клеток утолщены неравномерно: стенки, направленные к щели (брюшные), значительно утолщены по сравнению со стенками, направленными от щели (спинными). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью расположена крупная дыхательная полость (межклетник), окруженная клетками мезофилла листа.
Замыкающие клетки окружены околоустьичными
клетками, которые совместно
образуют устьичный комплекс
(рис.
2.7). Различают следующие основные типы
устьичных комплексов: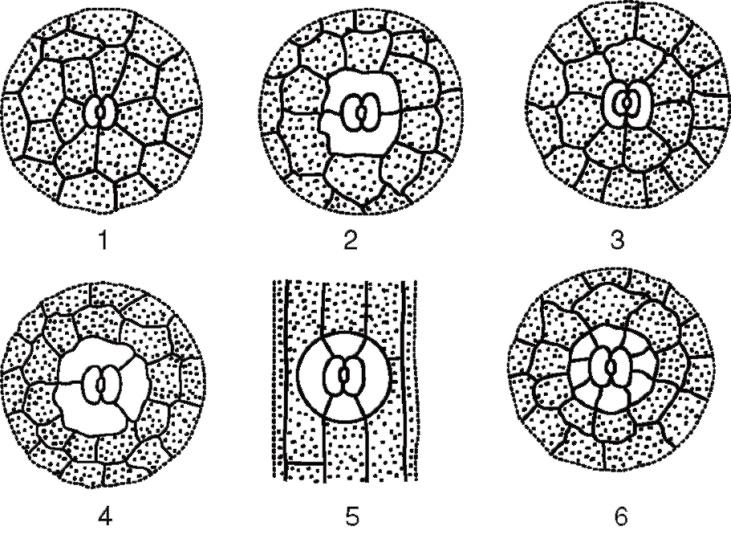
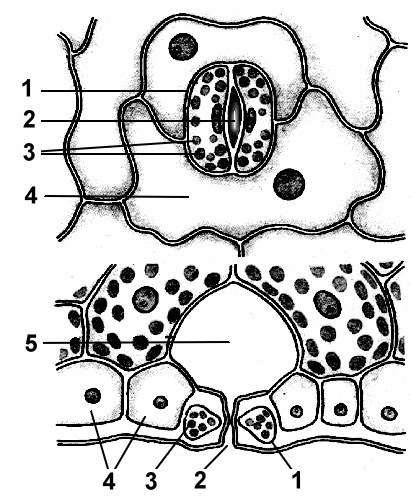
Рис. 2.7. Основные типы устьичного аппарата: 1 - аномоцитный (у всех высших растений, кроме хвощей); 2 - диацитный (у папоротников и цветковых); 3 - парацитный (у папоротников, хвощей, цветковых и гнетовых); 4 - анизоцитный (только у цветковых); 5 - тетрацитный (главным образом у однодольных); 6 - анциклоцитный (у папоротников, голосеменных и цветковых)
1) аномоцитный (беспорядочный) - замыкающие клетки не имеют ярко выраженных околоустьичных клеток; характерен для всех высших растений, исключая хвойные;
2) анизоцитный (неравноклеточный) - замыкающие клетки устьица окружены тремя околоустьичными клетками, одна из которых намного крупнее (или меньше) остальных;
3) парацитный (параллельно-клеточный) - одна околоустьичная клетка (или более) расположена параллельно замыкающим;
4) диацитный (перекрестно-клеточный) - две околоустьичные клетки расположены перпендикулярно замыкающим;
5) тетрацитный (от греч.tetra - четыре) - главным образом у однодольных;
Устьица расположены на нижней стороне
листа, но у водных растений с плавающими
листьями они находятся только на верхней
стороне листа. По форме клеток эпидермы
листа и расположению устьиц можно
отличить однодольное растение от
двудольного (рис. 2.8). Собственно
эпидермальные клетки листа двудольных
растений в очертаниях волнистые (рис.
2.9), а у однодольных они вытянутые,
ромбической формы.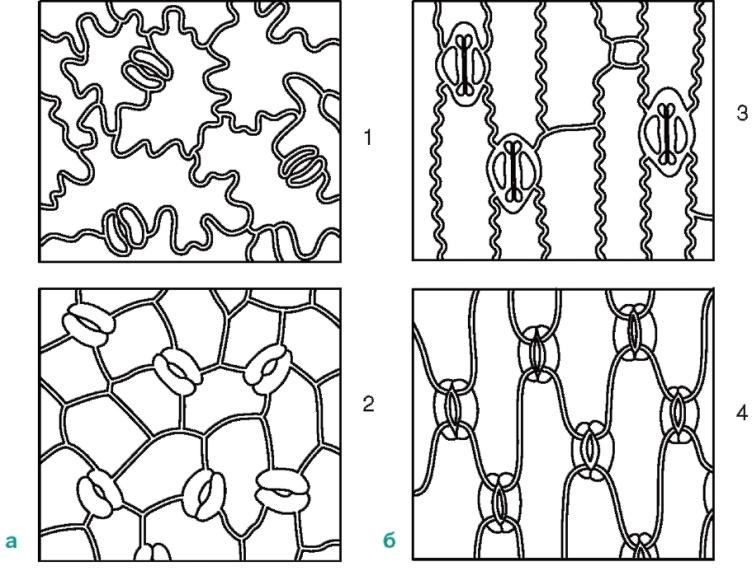
Рис. 2.8. Расположение устьиц на эпидерме (вид с поверхности):а -двудольные растения: 1 - буквица; 2 - арбуз;б -однодольные растения: 3 - кукуруза; 4 - ирис
Типы устьиц по уровню расположения относительно поверхности эпидермиса можно подразделить следующим образом.
1.7.1. Устьица, расположенные в одной плоскости с эпидермисом. Наиболее распространенный тип и обычно в описании микроскопии лекарственного растительного сырья не указывается, т.е. данный пункт опускается. Диагностическими признаками будут либо выступающие, либо погруженные устьица.
1.7.2. Выступающие устьица - устьица, расположенные над эпидермисом. Обычно при вращении микровинта микроскопа (при опускании объектива) сначала обнаруживаются такие устьица, а уже затем появляются клетки эпидермиса, поэтому на фотографии запечатлеть их с поверхности листа практически невозможно, равно как и изобразить на рисунке. В одной плоскости с эпидермисом такие устьица можно увидеть на поперечных срезах, но для этого срез должен пройти через устьице, что трудно получить при их редком расположении на листе. Такие устьица характерны, например, для листьев толокнянки.
1.7.3. Погруженные устьица - устьица, погруженные в эпидермис. При наблюдении под микроскопом при вращении микровинта (при опускании объектива) сначала четко обнаруживаются клетки эпидермиса, затем становится возможным более четко увидеть контуры устьиц. На фотографиях и рисунках препаратов с поверхности их также трудно отобразить. Встречаются в листьях ландыша, листьях вахты, листьях эвкалипта. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками и называютсяустьичными криптами.
1.8. Типы устьичных клеток
В литературе описывается 19 типов , нами выбраны только те, которые используются в анализе лекарственного растительного сырья**.
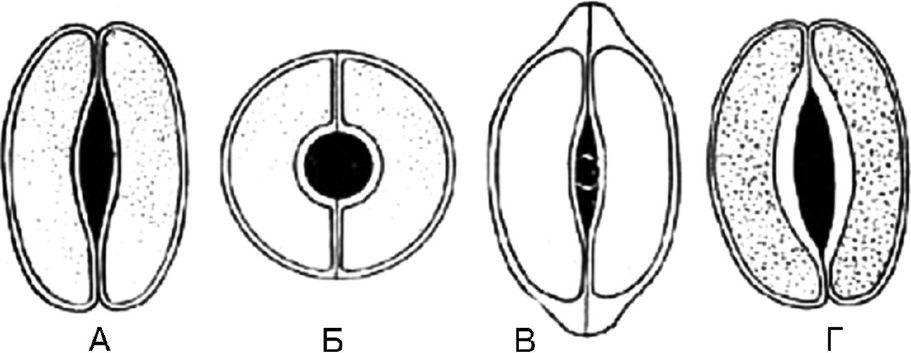
Рис. 63. Типы устьичных клеток. А - чечевицевидные; Б - сферовидные; В - колпачковидные; Г – ладьевидные
1.8.1. Чечевицевидные - 2 одинаковые клетки полулунной формы расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель веретеновидная (рис. 63, А). Тип устьичных клеток характерен для большинства растений.
1.8.2. Сферовидные - две одинаковые, сильно кругообразноизогнутые клетки расположены симметрично. На фронтальной плоскости утолщение оболочки почти равномерное. Щель круглая (рис. 63, Б).
1.8.3. Колпачковидные - две одинаковые клетки полулунной формы в полярных частях имеют утолщения в виде колпачка. Щель веретеновидная (рис. 63, В). Встречаются у наперстянок.
1.8.4. Ладьевидные - внутренние стенки устьичных клеток утолщены. Щель веретеновидная (рис. 63, Г). Наблюдается в траве золототысячника, в листьях вахты.
Механизм работы устьиц обусловлен осмотическими свойствами клеток. При освещении поверхности листа солнцем в хлоропластах замыкающих клеток происходит активный процесс фотосинтеза. Насыщение клеток продуктами фотосинтеза, сахарами влечет за собой активное поступление в клетки ионов калия, вследствие чего концентрация клеточного сока в замыкающих клетках возрастает. Возникает разность концентрации клеточного сока околоустьичных и замыкающих клеток. В силу осмотических свойств клеток вода из околоустьичных клеток поступает в замыкающие, что ведет к увеличению объема последних и резкому возрастанию тургора. Утолщение «брюшных» стенок замыкающих клеток, обращенных к устьичной щели, обеспечивает неравномерное растяжение клеточной стенки; замыкающие клетки приобретают выраженную бобовидную форму, и устьичная щель открывается. При снижении интенсивности фотосинтеза (например, вечером) уменьшается образование сахаров в замыкающих клетках. Приток ионов калия прекращается. Концентрация клеточного сока в замыкающих клетках снижается по сравнению с околоустьичными. Вода путем осмоса уходит из замыкающих клеток, понижая их тургор, в результате ночью устьичная щель закрывается.
Клетки эпидермы плотно сомкнуты между собой, благодаря этому эпидерма выполняет целый ряд функций:
Препятствует проникновению болезнетворных организмов внутрь растения;
Защищает внутренние ткани от механических повреждений;
Регулирует газообмен и транспирацию;
Через нее выделяются вода, соли;
Может функционировать как всасывающая ткань;
принимает участие в синтезе различных веществ, восприятии раздражений и в движении листьев.
Трихомы - различные по форме, строению и функциям выросты клеток эпидермы: волоски, чешуйки, щетинки и т.п. Их подразделяют на кроющие и железистые.Железистые трихомы, в отличие от кроющих, имеют клетки, выделяющие секрет.Кроющие волоски, образуя на растении шерстистый, войлочный или иной покров, отражают часть солнечных лучей и тем самым уменьшают транспирацию. Иногда волоски находятся только там, где расположены устьица, например, на нижней стороне листа мать-и-мачехи. У некоторых растений живые волоски увеличивают общую испаряющую поверхность, что способствует ускорению транспирации.
Размеры трихом значительно варьируют. Наиболее длинные трихомы (до 5-6 см) покрывают семена хлопчатника. Кроющие трихомы имеют форму простых одно или многоклеточных, разветвленных или звездчатых волосков. Кроющие трихомы могут длительное время оставаться живыми или быстро отмирать, заполняясь воздухом.
От трихом, возникающих только при участии эпидермальных клеток, отличаются эмергенцы, в образовании которых участвуют и более глубоко расположенные ткани субэпидермальных слоев.
Анатомо-диагностические признаки, имеющие наибольшее значение и высокую вариабельность при определении лекарственного сырья. Волоски могут быть простые и головчатые, которые в свою очередь могут быть одноклеточными и многоклеточными. Многоклеточные волоски могут быть однорядными, двухрядными и ветвистыми.


Устьица - это отверстия в эпидермисе, через которые происходит газообмен. Они находятся в основном на листьях, но имеются также и на стеблях. Каждое устьице окружено двумя замыкающими клетками, которые в отличие от обычных эпидермальных клеток содержат хлоропла-сты. Замыкающие клетки контролируют величину отверстия устьица за счет изменения своей тургесцентности. Внешний вид устьиц и замыкающих клеток хорошо видны на микрофотографиях, полученных с помощью сканирующего электронного микроскопа.
В статье мы уже говорили, как выглядят клетки эпидермиса, замыкающие клетки и устьица , если их рассматривать сверху в световом микроскопе. На рисунке приведено схематическое изображение устьица в разрезе. Видно, что стенки замыкающих клеток неравномерно утолщены: стенка, которая ближе к отверстию устьица, называемая вентральной, толще, чем противоположная, называемая дорсальной. Кроме того, целлюлозные микрофибриллы в стенке ориентированы таким образом, что вентральная стенка менее эластична, чем дорсальная. Некоторые микрофибриллы образуют как бы обручи вокруг замыкающих клеток, похожих на сардельки.
Эти обручи не эластичны, и по мере заполнения клетки водой,т. е. роста ее тургора, они не дают увеличиваться ее диаметру, позволяя растягиваться только в длину. Но поскольку замыкающие клетки соединены своими концами, а тонкие дорсальные стенки растягиваются легче, чем толстые вентральные, клетки приобретают полукруглую форму. В результате между двумя соседними замыкающими клетками появляется зазор, называемый устьичной щелью. Такой же эффект наблюдается, если надувать два скрепленных концами продолговатых воздушных шарика, наклеив вдоль их соприкасающихся сторон липкую ленту (имитация нерастяжимой вентральной стенки). Для полноты картины можно неплотно обмотать их такой же лентой по спирали, имитировав целлюлозные обручи.
Когда замыкающие клетки теряют воду и тургор, устьичная щель закрывается. Каким образом происходит изменение тургесцентности клеток, пока не ясно.
Согласно классической, так называемой «сахаро-крахмально й» гипотезе, в светлое время суток в замыкающих клетках повышается концентрация водорастворимых Сахаров, а следовательно, осмотический потенциал их становится более отрицательным, что стимулирует поступление в них воды путем осмоса. Однако никому еще не удалось показать, что в замыкающих клетках накапливается достаточное количество сахара, чтобы вызвать наблюдаемые изменения осмотического потенциала.
Недавно было установлено, что днем на свету в замыкающих клетках интенсивно накапливаются катионы калия и сопутствующие им анионы: они и играют роль, отводившуюся раньше сахару. До сих пор неясно, уравновешиваются ли при этом их заряды. У некоторых изученных растений отмечалось накопление на свету большого количества анионов органических кислот, в частности малата. Вместе с тем размеры крахмальных зерен, появляющихся в темноте в хлоропластах замыкающих клеток, уменьшаются. Дело в том, что крахмал на свету (необходимы синие лучи спектра) превращается в малат, возможно, по следующей схеме:
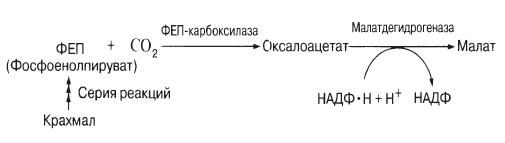
У некоторых видов, например у лука, крахмала в замыкающих клетках нет. Поэтому при раскрытых устьицах малат не накапливается, а катионы, по-видимому, поглощаются вместе с неорганическими ионами типа хлорид-ионов.
В темноте калий (К+) выходит из замыкающих клеток в окружающие эпидермальные клетки . В результате водный потенциал замыкающих клеток растет, и вода из них устремляется туда, где он ниже. Тургор замыкающих клеток падает, они изменяют форму, и устьичная щель закрывается.
Некоторые вопросы пока остаются без ответа. Например, почему на ceeiy в замыкающие клетки поступает калий? Какова роль хлоропластов, кроме накопления крахмала? Возможно, калий поступает внутрь благодаря «включению» АТФ-азы, локализованной в плазмалемме. Судя по некоторым данным, этот фермент активируется синим светом. Возможно, АТФаза нужна для откачивания из клетки протонов (Н+), а катионы калия движутся в клетку для уравновешивания заряда (аналогичный, обсуждаемый в разд. 13.8.4 насос работает во флоэме). Действительно, как и предполагает эта гипотеза, внутри замыкающих клеток рН на свету падает. В 1979 г. было показано, что в хлоропластах замыкающих клеток конских бобов (Vtcia faba) нет ферментов цикла Кальвина, а тилакоидная система развита слабо, хотя хлорофилл там имеется. Следовательно, обычный Сз-фотосинтез не работает, крахмала таким путем не образуется. Это, вероятно, объясняет, почему крахмал образуется не днем, как в обычных фотосинтезирующих клетках, а ночью.
Клетки эпидермиса почти непроницаемы для воды и газов благодаря своеоб-разному устройству их наружной стенки. Как же осуществляются газообмен между растением и наружной средой и испарение воды — процессы, не-обходимые для нормальной жизнедеятельности растения? Среди клеток эпидермиса встречаются характерные образования, называемые устьи-цами .
Устьице — щелевидное отверстие, окаймленное с двух сторон двумя замыкающими клетками, имеющими большей частью полулун-ную форму.
Клетки эти живые и со-держат хлорофилловые зерна и кру-пинки крахмала, отсутствующие в других клетках эпидермиса. Особен-но много устьиц на листе. Вид усть-ица с поверхности и в разрезе изоб-ражен на рисунке 40. На поперечном разрезе видно, что непосредственно под устьицем внутри ткани листа находится полость, называемая дыхательной . В пределах щели за-мыкающие клетки более сближены в средней части клеток, а выше и ниже они дальше отступают друг от друга, образуя пространства, называемые передним и задним дво-риком .
Замыкающие клетки способны увеличивать и сокращать свои разме-ры, благодаря чему устьичная щель то широко раскрывается, то сужива-ется или даже совсем бывает закрыта.
Таким образом, замыкающие клетки являются аппаратом, регулирующим процесс открывания и закрывания устьиц.
Как же осуществляется этот процесс?
Стенки замыкающих клеток, обращенные к щели, утолщены значи-тельно сильнее, чем стенки, обращенные к соседним клеткам эпидер-миса. Когда растение освещено и имеет избыток влаги, в хлорофилловых зернах замыкающих клеток происходит накопление крахмала, часть ко-торого превращается в сахар. Сахар, растворенный в клеточном соке, при-тягивает воду из соседних клеток эпидермиса, вследствие чего в замы-кающих клетках повышается тургор. Сильное давление приводит к выпячиванию стенок клеток, примыкающих к эпидермальным, а про-тивоположные, сильно утолщенные стенки выпрямляются. Вследствие этого устьичная щель раскрывается, и газообмен, а также испарение воды увеличиваются. В темноте или при недостатке влаги тур-горное давление уменьшается, замыкающие клетки принимают прежнее по-ложение и утолщенные стенки смыкаются. Щель устьица закрывается.
Устьица расположены на всех молодых неодревесневших наземных ор-ганах растения. Особенно много их на листьях, причем здесь они располо-жены, главным образом, на нижней поверхности. Если лист расположен вертикально, то устьица развиваются с обеих его сторон. У плавающих на поверхности воды листьев некоторых водных растений (например, кувшинки, кубышки) устьица расположены только на верхней стороне листа. Материал с сайта
Число устьиц на 1 кв. мм листовой поверхности в среднем равно 300, однако иногда достигает 600 и более. У рогоза (Typha ) насчитывают свыше 1300 устьиц на 1 кв. мм. Листья, погруженные в воду, устьиц не имеют. Расположены устьица чаще всего равномерно по всей поверхности кожи-цы, но у некоторых растений собраны группами. У однодольных растений, а также на хвоинках многих хвойных они расположены продольными рядами. У растений засушливых областей неред-ко устьица бывают погружены в ткань листа.
Развитие устьиц обычно происхо-дит следующим образом. В отдельных клетках эпидермиса образуются дуго-образные стенки, разделяющие клетку на несколько более мелких так, что центральная из них становится родо-начальницей устьиц. Эта клетка раз-деляется продольной (по оси клетки) перегородкой. Затем эта перегородка расщепляется, и образуется щель. Ог-раничивающие ее клетки становятся замыкающими клетками устьица. У не-которых печеночных мхов имеются свое-образные устьица, лишенные замыкаю-щих клеток.
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги , который это открытие опубликовал в 1675 году в своей работе Anatome plantarum . Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых . Прогресс в изучении наступил в XIX веке , и тогда же, в 1827 году , швейцарским ботаником Декандолем было впервые использовано слово „stoma“. Изучением устьиц в то время занимались Гуго фон Моль , который открыл основной принцип открывания устьиц и Симон Швенденер , классифицировавший устьица по типу их конструкции.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная (Commelina communis ), Боб садовый (Vicia faba ), Кукуруза сахарная (Zea mays ).
Строение
Размеры устьица (длина) колеблются в пределах 0,01-0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum , 0,12 мм (120 мкм) Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes) , которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis) . Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель . Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен . Воздух, содержащий диоксид углерода (углекислый газ) и кислород , проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса , примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями ) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный - сопровождающие клетки не отличаются от остальных клеток эпидермиса , тип весьма обычен для всех групп высших растений , за исключением хвойных ;
- диацитный - характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный - сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный - замыкающие клетки окружены тремя сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный - четыре сопровождающие клетки, характерен для однодольных ;
- энциклоцитный - сопровождающие клетки образуют узкое колесо вокруг замыкающих клеток;
- актиноцитный - несколько сопровождающих клеток, радиально расходящихся от замыкающих клеток;
- перицитный - замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный - замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный - замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный - устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный - такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
У двудольных распространённым является парацитный тип устьиц. Замыкающие клетки почковидной (бобовидной) формы - такими они видны с поверхности листа - несут хлоропласты , тонкие неутолщённые участки оболочки образуют выступы (носики) закрывающие устьичную щель.
Наружные стенки замыкающих клеток обычно имеют выросты, что хорошо видно на поперечном разрезе устьица. Пространство, ограниченное этими выростами, называют передним двориком. Нередко аналогичные выросты наблюдаются и у внутренних оболочек замыкающих клеток. Они образуют задний дворик, или внутренний, соединённый с крупным межклетником - подустьичной полостью.
У однодольных парацитное строение устьиц отмечено у злаковых . Замыкающие клетки имеют гантелевидную форму - сужены в средней части и расширены на обоих концах, при этом стенки расширенных участков очень тонкие, а в средней части замыкающих клеток сильно утолщены. Хлоропласты располагаются в пузыревидных окончаниях клеток.
Движение замыкающих клеток
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при неодинаковом водоснабжении в ночные часы, а иногда и днём тургор в замыкающих клетках понижается, и устьичная щель замыкается, снижая тем самым уровень транспирации . С повышением тургора устьица открываются. Считают, что главная роль в изменении тургора принадлежит ионам калия . Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар , повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и повышению тургорного давления в замыкающих клетках.
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обоих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу . Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды зимой на испарение, а летом - во время засухи.
У мхов (исключение антоцеротовые)настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.


