Поперечный разрез корня в зоне всасывания. Строение корня растения
В зоне деления корня в апикальной меристеме в определенной последовательности и строго закономерно возникают внутренние ткани. Причем, здесь есть четкое разделение на два отдела. От среднего слоя инициальных клеток происходит наружный отдел, который называется периблемой . От верхнего слоя инициальных клеток происходит внутренний отдел, его называют плеромой .
Из плеромы в последствии формируется стела (центральный цилиндр ), одни из ее клеток превращаются в сосуды и трахеиды, из других происходят ситовидные трубки, из третьих - клетки сердцевины и т.д.
Из клеток периблемы образуется первичная кора корня , которая состоит из паренхимных клеток основной ткани.
Из дерматогены (наружного слоя клеток), расположенной на поверхности корня, обособляется первичная покровная ткань, которую называют эпиблемой или ризодермой . Ризодерма — однослойная ткань, которая достигает своего полного развития в зоне поглощения.
является результатом дифференциации меристемы апекса. В первичной структуре корня в области его кончика, можно выделить 3 слоя: наружный — эпиблему , средний — первичную кору и центральный осевой цилиндр — стелу . См. рисунок ниже.

В сформированной ризодерме образуется множество тончайших выростов - корневых волосков (см. рисунки ниже).
Корневые волоски недолговечны. Воду и и растворённые в воде вещества они могут активно поглощать лишь только в растущем состоянии. Благодаря образованию волосков увеличивается более чем в 10 раз общая поверхность зоны всасывания. Как правило, длина волосков составляет не более 1 мм. Они покрыты очень тонкой оболочкой, состоящей из целлюлозы и пектиновых веществ.
В клетки корневых волосков вода проникает пассивно , а именно, благодаря разности в осмотическом давлении почвенного раствора и клеточного сока. А вот минеральные вещества поступают в корневые волоски в результате активного всасывания . Это процесс протекает с затратами энергии, чтобы преодолеть градиент концентрации. После попадания в цитоплазму, минеральные вещества передаются от корневого волоска до ксилемы от клетки к клетке. Благодаря корневому давлению, которое создается силой всасывания всех корневых волосков, а также испарению воды с поверхности листьев растения (транспирацией) обеспечивается движение почвенного раствора вверх по сосудам корня и стебля.
Все эти энергоемкие процессы растение может обеспечивать за счет дыхания !
В результате диффузии кислорода из почвы в ткани происходит дыхание. Для дыхания растениям необходимы органические вещества. Эти органические вещества поступают в корень из листьев. Энергия, образуемая в процессе дыхания, запасается в молекулах АТФ. Эта энергия будет расходоваться на деление клеток, рост, процессы синтеза, транспорт веществ и т.п. Именно по этой причине необходимо, чтобы в почву проникал воздух, а для этого почву надо рыхлить. Кроме того, благодаря рыхлению почвы в ней сохраняется влага, поэтому рыхление часто называют еще «сухим поливом».
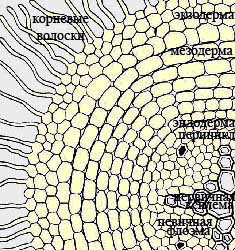
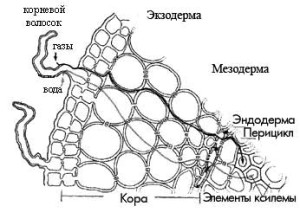
Первичная кора, которая, как было сказано выше, образуется из периблемы, состоит из живых тонкостенных паренхимных клеток. В первичной коре можно выделить 3 четко различающихся друг от друга слоя: эндодерму , мезодерму и экзодерму .
Эндодерма — это внутренний слой первичной коры, который прилегает непосредственно к центральному цилиндру или стеле. Эндодерма состоит из одного ряда клеток, у которых есть утолщения на радиальных стенках (также они называются пояски Каспари), чередуемых с тонкостенными пропускными клетками. Эндодерма контролирует прохождение веществ из коры в центральный цилиндр и обратно, так называемые горизонтальные токи.
Следующим слоем, идущим после эндодермы является мезодерма или средний слой первичной коры. В состав мезодермы входят клетки с системой межклетников, расположенные рыхло. По этим клеткам идет интенсивный газообмен. В мезодерме происходит синтез пластических веществ и дальнейшее их передвижение в другие ткани, накапление запасных веществ, а также располагается микориза.
Последний, наружный слой первичной коры называют экзодермой . Экзодерма располагается непосредственно под ризодермой, а по мере того, как отмирают корневые волоски, оказывается на поверхности корня. В данном случае экзодерма может выполнять функции покровной ткани: у нее происходит утолщение и опробковение клеточных оболочек, отмирание содержимого клеток. Среди этих опробковевших клеток остаются неопробковевшие пропускные клетки. Через эти пропускные клетки происходит прохождение веществ.
Наружный слой стелы, который примыкает к эндодерме, называют перициклом . Его клетки в течение длительного времени сохраняют способность к делению. В этом слое происходит зарожение боковых корешков, поэтому перицикл еще называют корнеродным слоем. Характерной чертой корней является чередование в стеле участков ксилемы и флоэмы. Ксилема образует звезду. У различных групп растений число лучей этой звезды может быть разным. Между лучами этой зведы располагается флоэма. В самом центре корня могут располагаться элементы первичной ксилемы, склеренхима или тонкостенная паренхима. Характерной особенностью корня, которая отличает его по анатомической структуре от стебля, является чередование первичной ксилемы и первичной флоэмы по периферии стелы.
Такое первичное строение корня характерно для молодых корней у всех групп высших растений. У папоротников, хвощей, плаунов и представителей класса однодольных цветковых растений первичная структура корня сохранятся в течение всей его жизни.
Вторичное строение корня.
У голосеменных и двудольных покрытосеменных растений первичная структура корня сохраняется только до начала процесса его утолщения Этот процесс — результат деятельности вторичных боковых меристем - камбия и феллогена (или пробкового камбия).
Началом процесса вторичных изменений является появление прослоек камбия под участками первичной флоэмы, направленных вовнутрь от неё. Возникает камбий из слабо дифференцированной паренхимы центрального цилиндра. Наружу он откладывает элементы вторичной флоэмы (или луба), а вовнутрь — элементы вторичной ксилемы (или древесины). В начале этого процесса прослойки камбия разобщены, в дальнейшем происходит их смыкание и образуется сплошной слой. Это происходит благодаря тому, что клетки перицикла интенсивно делятся напротив лучей ксилемы. Из камбиальных участков, которые возникли из перицикла, образуются только паренхимные клетки, так называемых сердцевинных лучей. А вот остальные клетки камбия образуют проводящие элементы: ксилему и флоэму.
На продольном разрезе через кончик корня можно обнаружить следующие участки: корневой чехлик , зону ро-ста , зону корневых волосков , зону со строением взрослого корня (рис. 59).
Зона роста расположена несколько отступя от кончика корня. Она составле-на несколькими рядами узких таблитчатых клеток с тонкими оболочками и протоплазмой, заполняющей полость клетки. Это образовательная ткань — меристема . Фактически имеются две меристемы: одна — кончика кор-ня, другая — корневого чехлика.
Клетки последней отчленяют новые клет-ки кнаружи и формируют корневой чехлик . Самые наружные клетки округ-ляются, разъединяются и отшелушиваются под механическим воздействием твердых частиц почвы. Отмершие клетки замещаются клетками, располо-женными ниже их и возникающими из меристемы корневого чехлика. Кнутри из меристемы кончика корня путем деления откладываются клетки собственно корня, из которых по мере дальнейшего развития и роста фор-мируются различные ткани корня. Несколько выше меристемы кончика корня на продольном разрезе уже заметна дифференцировка клеток кор-ня на 3 слоя:
- наружный слой, в дальнейшем развивающийся в кожицу — эпидермис корня;
- средний слой, дающий далее начало прово-дящей части корня;
- промежуточный слой, дальше развивающий так называемую кору корня (рис. 60 в, д, г).
По мере дальнейшего разви-тия эти три зоны внутреннего строения корня хорошо дифференцируются. На поперечном разрезе, проведенном в области развития корневых волосков, корень имеет строение, изображенное на рисунке 61.
Снаружи корень покрыт однослойным эпидермисом . Клетки эпидер-миса живые, имеют тонкие стенки. На периферии они вытянуты в длинные выросты — корневые волоски (рис. 61, а). Ядро клеток перемещается в корневой волосок, располагаясь в переднем, растущем его конце, на ниж-ней стороне (рис. 62).
Корневые волоски имеют длину 0,15—2,5 мм (во влажном воздухе и в искусственной обстановке могут достигать 8 мм). Число их на корне очень велико — в среднем на 1 лш 8 до 425 штук. Зона корневых волосков зани-мает участок корня длиной 1—2 см. Уже на расстоянии 3—8 мм от начала зоны корневые волоски отмирают, отпадают и по мере роста корня ближе к верхушке корня возникают все новые и новые. Корневые волоски выде-ляют наружу различные вещества (кислоты), которые растворяют нераство-римые в воде соединения, имеющиеся в почве, и делают их доступными для растений. Проникая между частицами почвы, корневые волоски плотно срастаются с ними. При выдергивании из земли и при дальнейшей пересад-ке растение часто страдает или даже совсем не приживается вследст-вие отрывания молодых, растущих частей корня с корневыми волос-ками.
За зоной корневых волосков расположена кора корня (рис. 61, к). Она представлена несколькими рядами многогранных паренхимных кле-ток. Самый наружный слой коры корня составлен более крупными, несколь-ко радиально вытянутыми клетками, плотно примыкающими к эпидермису. По опадении корневых волосков оболочки клеток этого слоя опробкове-вают; кроме того, под ними возникает слой пробковых клеток , которые, таким образом, защищают наружный слой корня от испарения. Глубоко лежащие слои корня состоят из более мелких клеток.
Самый внутренний слой коры образует так называемую эндодерму , отчленяющую кору от срединной части корня, так называемого центрального цилиндра , или стели (рис. 61, эн). Клетки эндо-дермы расположены в один слой и тесно придвинуты друг к другу. На-ружные стенки их тонкие, внутренние же и радиальные (боковые) стенки получают сильное утолщение и, пропитываясь суберином, опробковева-ют, становясь непроницаемыми для воды и газов. Среди клеток эндодермы встречаются отдельные клетки с неутолщенными стенками, богатые содер-жимым (пропускные клетки ); через эти клетки происходит про-никновение воды из коровой части корня в центральный цилиндр.
Центральный цилиндр, занимая срединную часть корня, состоит из различных тканей. Самый наруж-ный слой (реже два слоя) клеток, примыкающий изнутри к эндодер-ме, носит название перицикла (рис. 61). Клетки его более крупные, слегка вытянуты по радиусу и имеют тонкие клетчатковые оболочки. В этом слое про-исходит заложение боковых ко-решков, почему перицикл часто называют корнеродным слоем . Образуются они следую-щим образом: над группой сосу-дов в перицикле наблюдается уси-ленное деление его клеток, бла-годаря чему образуется небольшой бугорок, выпяченный в перифери-ческую сторону корня. Снаружи его облегают клетки эндодермы.
Постепенно в бугорке формируются слои, свойственные меристеме глав-ного корня, из которых в дальнейшем образуются соответственные ткани. Бугорок продолжает расти, окруженный слоем эндодермы (так называемый «кармашек»), и, прорывая кору, выходит наружу. Материал с сайта
Таким образом, боковые корни, в отличие от главного корня и стебля, возникают эндогенно (внутриродно). Эндогенно возникают все при-даточные корни, из какой бы части растения они ни образовались.
За перициклом расположена паренхимная ткань, в которую погружены проводящие элементы (сосудистые пучки ). Сосудистый пучок корня построен по радиальному типу (радиальный пучок). Ксилема его расположена группами, лучами, идущими от периферии к центру. Число лучей обычно небольшое — 3, 4, 5. Иногда группы ксилемы не доходят до центра, который заполнен тогда паренхимными клетками. Сосуды ксилемных лучей имеют более узкие просветы ближе к периферии и более крупные к центру. Наружные сосуды закладываются раньше цен-тральных. Развитие ксилемы в корне идет от периферии к центру. Наружные сосуды обычно бывают кольчатыми или спиральными; более крупные вну-тренние — пористые. Между лучами ксилемы в верхней части расположе-ны участки флоэмы, составленной ситовидными трубками с сопровождаю-щими клетками и лубяной паренхимой.
На этой странице материал по темам:
По происхождению корни делятся на главные , боковые и придаточные. Главный корень семенных растений развивается из корешка зародыша семени. Стебель является продолжением корня, и вместе они составляют ось 1-го порядка. Место сочленения оси и семядольных листьев называют семядольным узлом. Участок, расположенный на границе главного корня и стебля, именуется корневой шейкой. Участок стебля от корневой шейки до первых зародышевых листьев (семядолей) называют подсемядольным коленом , или гипокотилем, а от семядолей до первых настоящих листьев - эпикотилем , или надсемядольным коленом . У двудольных и голосеменных растений от главного корня за счет меристематической активности перицикла отходят боковые корни 1-го порядка, которые дают начало боковым корням 2-го и 3-го порядка. Корневая система, образованная системой главного корня, называется стержневой, а с развитой системой боковых корней - ветвистой; таким образом, ветвистая корневая система является разновидностью стержневой. Чем больше боковых корней отходит от главного, тем больше площадь питания растения.
У большинства двудольных растений главный корень сохраняется всю жизнь, у однодольных растений главный корень не развивается, так как зародышевый корешок быстро отмирает, а от базальной части побега берут начало придаточные корни. Придаточные корни могут образовываться от листьев, стеблей, старых корней и даже от цветков по происхождению: А - система главного корня; Б, В - смешанная корневая система; Г - придаточная корневая система; 1 - главный корень; 2 - боковые корни; 3 - придаточные корни; 4 - основания побегов и иметь ответвления 1-го, 2-го порядка и т.д. Корневая система, образованная придаточными корнями, называется мочковатой (см. рис. 3.2). У многих двудольных корневищных растений главный корень часто отмирает и преобладает система придаточных корней, отходящих от корневища (лютик ползучий, сныть обыкновенная).
Рис. 3.2. Типы корневых систем: по форме:
А, Б - стержневая; В, Г - мочковатая;
по происхождению: А - система главного корня; Б, В - смешанная корневая система; Г - придаточная корневая система; 1 - главный корень; 2 - боковые корни; 3 - придаточные корни; 4 - основания побегов
По отношению к субстрату корни бывают следующих типов: земляные - развиваются в почве; водные - находятся в воде (у плавающих водных растений); воздушные, развивающиеся в воздушной среде (у растений, имеющих корни на стволах и листьях).
Зоны корня
В молодом корне различают 4 зоны: деления, растяжения, всасывания, проведения (см. рис. 3.3).
К зоне деления относят верхушку конуса нарастания (протяженность менее 1 мм), где происходит активное митотическое деление клеток. Верхушечная меристема откладывает наружу клетки корневого чехлика, а внутрь - ткани остальной части корня. Эта зона состоит из тонкостенных паренхимных клеток первичной меристемы, которые прикрыты корневым чехликом, выполняющим защитную функцию при продвижении корня между частичками почвы. От соприкосновения с почвой клетки чехлика постоянно разрушаются, образуя слизь, которая предохраняет зону деления при трении о
почву и продвижении корня вглубь. У большинства растений корневой чехлик восстанавливается за счет первичной меристемы, а у злаков - за счет особой меристемы калиптрогена.
По теории гистогенов (Ганштейн, 1868), у большинства покрытосеменных растений апикальные меристемы состоят из 3 гистогенных слоев, различающихся направлением деления клеток и имеющих по 1-4 инициальные клетки. Самый наружный слой - дерматоген - формирует протодерму, из которой образуются клетки корневого чехлика и ризодерма - первичная покровно-всасывающая ткань в зоне всасывания. Средний слой - периблема - дает начало всем тканям первичной коры. Третий слой инициалей формирует плером, из которого развиваются ткани центрального осевого цилиндра.
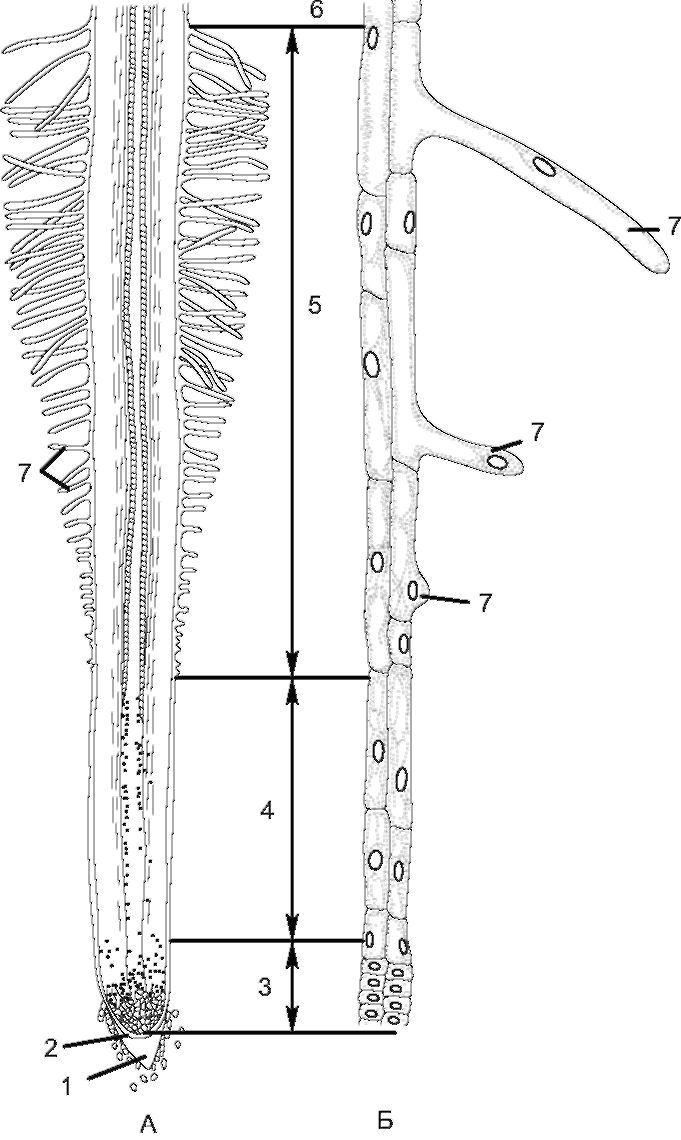
Рис. 3.3. Зоны корня (у проростка пшеницы):
А - схема строения корня; Б - периферические клетки отдельных зон при большом увеличении: 1 - корневой чехлик; 2 - калиптроген; 3 - зона деления; 4 - зона растяжения; 5 - зона всасывания; 6 - зона проведения;
7 - корневые волоски
В зоне растяжения клетки меристемы увеличиваются в размерах (вследствие оводнения), вытягиваются в длину, и деление клеток постепенно прекращается. Вследствие вытягивания клеток в продольном направлении происходят рост корня в длину и его продвижение в почве. Зону деления и зону растяжения с учетом сохранения в них меристематической активности можно объединить в одну - зону роста. Ее протяженность составляет несколько миллиметров. В зоне всасывания происходит становление первичной структуры корня.
Протяженность зоны всасывания - от нескольких миллиметров до нескольких сантиметров; она характеризуется наличием корневых волосков, являющихся выростами клеток ризодермы. При их формировании ядро перемещается в переднюю часть корневого волоска. Последние увеличивают всасывающую поверхность корня и обеспечивают активное всасывание воды и растворов солей, но они недолговечны (живут 10-20 дней). Новые корневые волоски образуются под зоной всасывания, а отмирают - над этой зоной. По мере роста растения зона всасывания постепенно перемещается, и растение имеет возможность поглощать минеральные вещества из разных слоев почвы.
Постепенно зона всасывания переходит в зону проведения (укрепления). Она тянется вплоть до корневой шейки и составляет по протяженности большую часть корня. В этой зоне идет интенсивное ветвление главного корня и появляются боковые корни. У двудольных растений в зоне проведения формируется вторичная структура корня.
Анатомия корня
Первичное строение корня (рис. 3, см. цв. вкл.). Строение корня в зоне всасывания называется первичным, потому что здесь происходит дифференциация тканей из первичной меристемы конуса нарастания. Первичное строение корня в зоне всасывания можно наблюдать у двудольных и однодольных растений, но у однодольных оно сохраняется на протяжении всей жизни растения. На поперечном срезе корня первичного строения выделяют 3 основные части: покровно-всасывающую ткань, первичную кору и центральный осевой цилиндр (рис. 3.4).
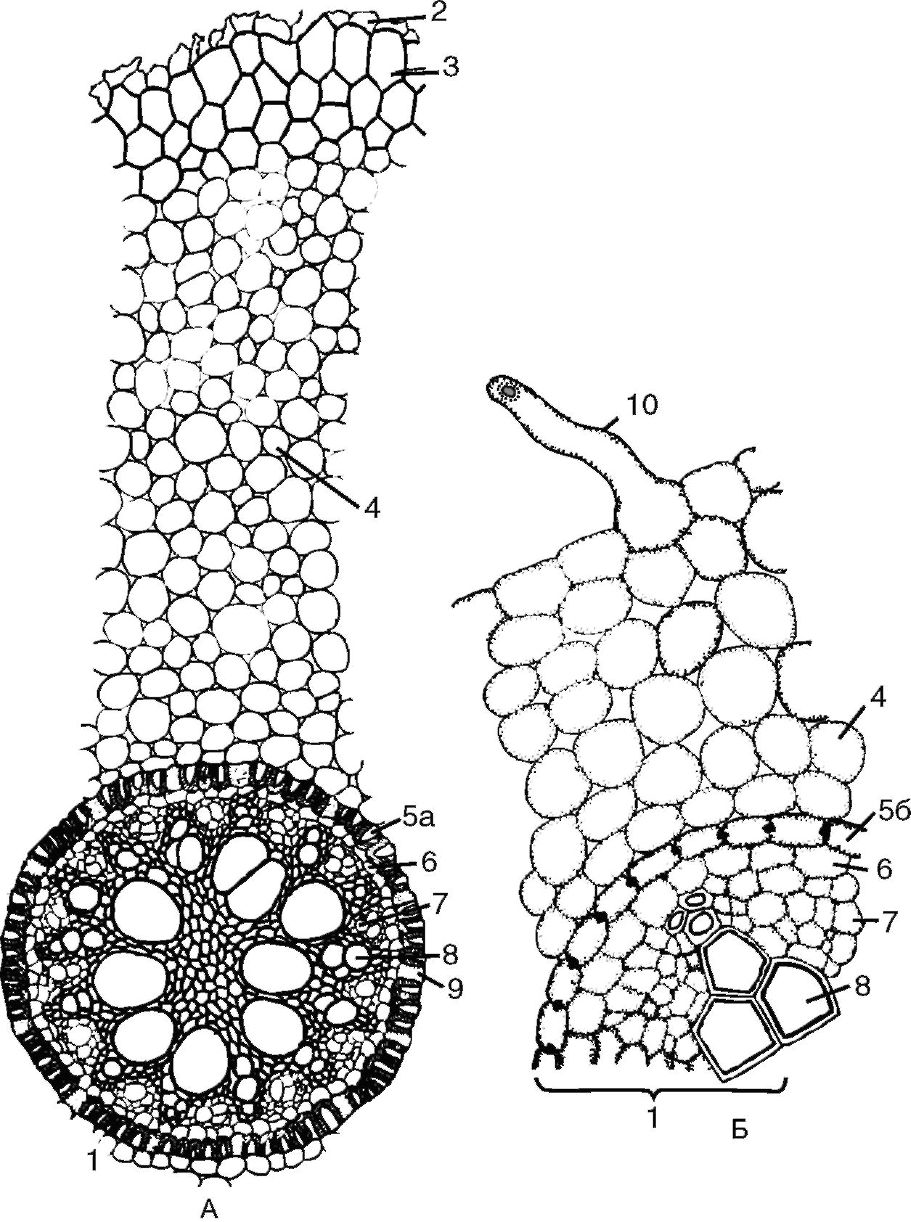
Рис. 3.4. Поперечный срез корня первичного строения:
А - первичное строение корня однодольного; Б - первичное строение корня двудольного: 1 - центральный (осевой) цилиндр; 2 - остатки эпиблемы; 3 - экзодерма; 4 - мезодерма; 5а - эндодерма с подковообразными утолщениями; 5б - эндодерма с поясками Каспари; 6 - перицикл; 7 - первичная флоэма; 8 - сосуды первичной ксилемы; 9 - пропускные клетки эндодермы; 10 - корневой волосок
Покровно-всасывающая ткань - ризодерма (эпиблема) выполняет как покровную функцию, так и функцию интенсивного всасывания воды и минеральных веществ из почвы. Клетки ризодермы живые, с тонкой целлюлозной стенкой. Из некоторых клеток ризодермы формируются корневые волоски; каждый из них представляет собой длинный вырост одной из клеток ризодермы, ядро же клетки обычно находится в кончике выроста. Корневой волосок содержит тонкий пристенный слой цитоплазмы, более плотный на верхушке волоска, а в центре - крупную вакуоль. Корневые волоски недолговечны и в зоне укрепления отмирают. Физиологически зона всасывания представляет собой очень важную часть корня. Клетки ризодермы поглощают водные растворы всей поверхностью наружных стенок. Развитие корневых волосков во много раз увеличивает поверхность поглощения. Протяженность зоны всасывания - от 1 до 1,5 см.
Со временем эпиблема может слущиваться, и тогда покровную функцию выполняет экзодерма, а после ее разрушения - слой кле- ток мезодермы и иногда мезодермы и перицикла, стенки которых опробковевают и одревесневают. Поэтому диаметр старых корней однодольных растений меньше, чем молодых.
Первичная кора корня развита более мощно, чем центральный осевой цилиндр. Она состоит из 3 слоев: экзодермы, мезодермы (рис. 4, см. цв. вкл.) (паренхима первичной коры) и эндодермы. Клетки экзодермы многоугольные по форме, плотно сомкнуты и расположены в несколько рядов. Клеточные стенки пропитаны суберином, т.е. опробковевают. Опробковение обеспечивает непроницаемость клеток для воды и газов. В экзодерме, обычно под корневыми волосками, сохраняются клетки с тонкими целлюлозными стенками - пропускные клетки, через которые проходят вода и минеральные вещества, поглощенные ризодермой. Обычно они располагаются напротив лучей ксилемы радиального пучка.
Под экзодермой находятся живые паренхимные клетки мезодермы . Это наиболее широкая часть первичной коры. Клетки мезодермы выполняют запасающую функцию, а также функцию проведения воды и растворенных в ней солей от корневых волосков в центральный осевой цилиндр.
Внутренний однорядный слой первичной коры представлен эндодермой. Клетки эндодермы плотно сомкнуты и почти квадратные в поперечном сечении. В зависимости от степени утолщения клеточной стенки различают 2 типа эндодермы - с поясками Каспари (на поперечном срезе они выглядят как пятна Каспари) и с подковообразными утолщениями стенок.
Эндодерма с поясками Каспари - это начальный этап формирования эндодермы, при котором утолщению подвергаются лишь ее радиальные стенки за счет отложения веществ, сходных по химическому составу с суберином и лигнином. У многих двудольных и голосеменных растений процесс дифференциации эндодермы поясками Каспари заканчивается. У эндодермы с подковообразными утолщениями образуется толстая вторичная клеточная стенка, пропитанная суберином, в дальнейшем она одревесневает. Неутолщенной остается только наружная клеточная стенка (рис. 3.5). Эндодерма с подковообразными утолщениями развивается чаще у однодольных растений (рис. 5, см. цв. вкл.).
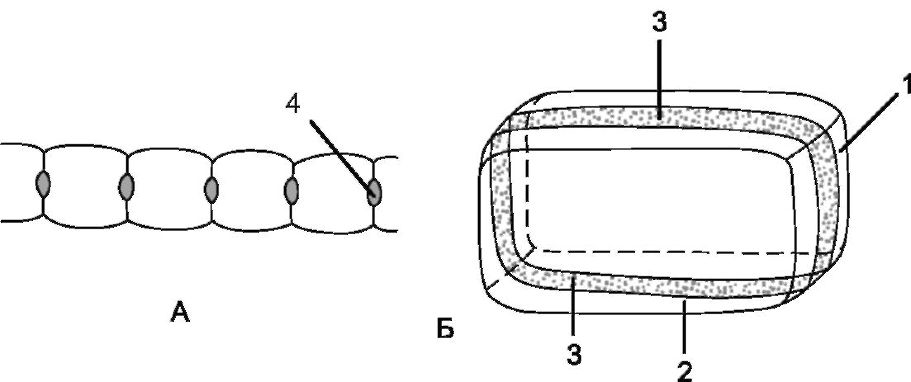
Рис. 3.5. Схема строения клетки эндодермы:
А - общий вид; Б - поперечный разрез клеток: 1 - поперечная стенка клетки; 2 - продольная радиальная стенка; 3 - поясок Каспари; 4 - пятна Каспари
Считается, что эндодерма выполняет функцию гидравлического барьера, способствуя продвижению минеральных веществ и воды из первичной коры в центральный осевой цилиндр и препятствуя их выходу обратно.
Центральный осевой цилиндр начинается с клеток перицикла, который обычно в молодых корнях состоит из живых тонкостенных паренхимных клеток, расположенных в один ряд (но может быть и многослойным - например, у грецкого ореха). Клетки перицикла дольше других тканей корня сохраняют свойства меристемы и спо- собность к новообразованиям. Из перицикла образуются боковые корни, поэтому его называют корнеродным слоем. Проводящая система корня представлена одним радиальным сосудисто-волокнистым пучком, в котором группы элементов первичной ксилемы чередуются с участками первичной флоэмы. У однодольных растений количество лучей первичной ксилемы - 6 и более, у двудольных - от 1 до 5. Корни в отличие от стеблей не имеют сердцевины, так как в центре корня располагаются лучи первичной ксилемы.
У однодольных и споровых архегониальных растений строение корня не претерпевает значительных изменений в течение всей жизни растения. У голосеменных и двудольных растений на границе зон всасывания и проведения происходит переход от первичного строения корня ко вторичному (табл. 3.1).
Основные функции корня: обеспечивает закрепление растения в почве, всасывание почвенного водного раствора солей и транспорт его к надземным частям растения.
Дополнительные функции: запасание питательных веществ, фотосинтез, дыхание, вегетативное размножение, выделение, симбиоз с микроорганизмами, грибами. Первые настоящие корни появились у папоротникообразных.
Зародыш корня называется зародышевым корнем и закладывается одновременно с почкой в зародыше семени.
У растений различают:
Главный корень. Он образуется из зародышевого и сохраняется на протяжении всей жизни. Всегда один.
Боковые корни. Ответвляются от корней (главного, дополнительных, боковых). Образуют при ветвлении корни 2-го, 3-го и т. д. порядка.
Придаточные корни. Образуются в любой части растения (стебле, листьях).
Совокупность всех корней растения образует корневую систему. Корневая система формируется в течение всей жизни растения. Ее формирование обеспечивают преимущественно боковые корни. Различают два типа корневой системы: стержневую и мочковатую.
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Меристемы- образовательные ткани- расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля).
Определение размеров корневых систем требует специальных методов. Очень много в этом отношении достигнуто благодаря работам русских физиологов В.Г. Ротмистрова, А.П. Модестова, И.В. Красовской. Оказалось, что общая поверхность корней обычно превышает поверхность надземных органов в 104-150 раз. При выращивании одиночного растения ржи было устоновленно, что общая длинна его корней достигает 600 км., при этом на них образуется 15 млрд. корневых волосков. Эти данные говорят об огромной потенциальной способности к росту корневых систем. Однако эта способность не всегда проявляется. При росте растений в фитоценозах, с достаточно большой густотой их строение, размеры корневых систем заметно уменьшаются.
С физиологической точки зрения корневая система не однородна. Довольно не вся поверхность корня участвует в поглощении волы. В каждом корне различают несколько зон (рис.1). Правда, не всегда все зоны выражены одинаково четко.
Окончание корня с наружи защищено корневым чехликом, напоминающим округлый колпачок, таящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика слущиваются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усилено делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона-зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки, затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а так же расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.
Рис. 1. Схема строения корня:
А - продольный разрез: 1-корневой чехлик; 2- меристема; 3-зона растяжения; 4- зона корневых волосков; 5- зона ветвления;
Б - поперечный разрез (по М.Ф.
Даниловой): 1 - ризодерма; 2 - корневой волосок; 3 - паренхима; 4 - эндодерма; 5-
пояски Каспари; 6 - перицикл; 7 - флоэма; 8 - ксилема. Пунктирные стрелки- пути
передвижения веществ, поглощаемых из наружного раствора. Сплошные стрелки путь
растворов по симпласту; прерывистые - путь по апопласту.
Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системных крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток- эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином. При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения, как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня.. При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление волосков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является основной зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет 5-10 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче.
Корень по длине можно разделить на несколько участков, имеющих различное строение и выполняющих различные функции. Эти участки называют зонами корня. Выделяют корневой чехлик и следующие зоны: деления, растяжения, всасывания и проведения.
Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы конуса нарастания. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует самый поверхностный слой с корневыми волосками - ризодерма (эпиблема). Защитную функцию выполняет ниже лежащая ткань - экзодерма.
В первичном строении корня выделяют три части: ризодерму, первичную кору и
осевой (центральный) цилиндр.
Строение ризодермы рассматривалось в теме "Покровные ткани".
На первичную кору приходится основная масса первичных тканей корня. Ее клетки
накапливают крахмал и другие вещества. Эта ткань содержит многочисленные
межклетники, имеющие значение для аэрации клеток корня. Наружные клетки
первичной коры, лежащие непосредственно под ризодермой, называются экзодермой.
Основная масса коры (мезодерма) образована паренхимными клетками. Самый
внутренний слой носит название эндодермы. Это ряд плотно сомкнутых клеток (без
межклетников).
Центральный или осевой цилиндр (стела) состоит из проводящих тканей, окруженных
одним или несколькими слоями клеток - перициклом.
Внутренняя часть центрального цилиндра у большинства растений занимает сплошной
тяж первичной ксилемы, дающий к перициклу выступы в виде ребер. Между ними
размещаются тяжи первичной флоэмы.
У двудольных и голосеменных растений уже в раннем возрасте в центральном цилиндре корня между ксилемой и флоэмой появляется камбий, деятельность которого приводит к вторичным изменениям и в конечном итоге формируется вторичная структура корня. К центру камбий откладывает клетки вторичной ксилемы, а к периферии - клетки вторичной флоэмы. В результате деятельности камбия первичная флоэма оттесняется кнаружи, а первичная ксилема остается в центре корня.
Вслед за изменениями в центральном цилиндре корня происходят изменения в коровой части. Клетки перицикла начинают делиться по всей окружности, в результате чего возникает слой клеток вторичной меристемы - феллогена (пробкового камбия). Феллоген, в свою очередь, делясь, откладывает наружу феллему, а внутрь - феллодерму. Образуется перидерма, пробковый слой которой изолирует первичную кору от центрального цилиндра. В результате вся первичная кора отмирает и постепенно сбрасывается; наружным слоем корня становится перидерма. Клетки феллодермы и остатки перицикла в дальнейшем разрастаются и составляют паренхимную зону, которую называют вторичной корой корня (рис. 2).
При развитии запасающей паренхимы главного корня происходит формирование запасающих корней или корнеплодов. Различают корнеплоды:
1. Монокамбиальные (редька, морковь) - закладывается только один слой камбия, а запасные вещества могут накапливаться либо в паренхиме ксилемы (ксилемный тип - редька), либо в паренхиме флоэмы (флоэмный тип - морковь);
2. Поликамбиальные - через определенные промежутки времени происходит заложение нового слоя камбия (свекла).
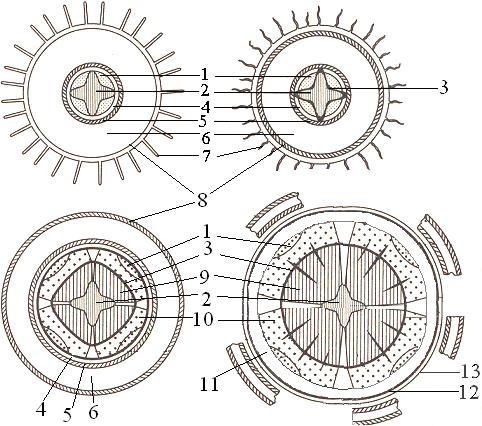
Рис. 2. Переход от первичного строения корня к вторичному:
1 - первичная флоэма, 2 - первичная ксилема, 3 - камбий, 4 - перицикл, 5 - эндодерма, 6 - мезодерма, 7 - ризодерма, 8 - экзодерма, 9 - вторичная ксилема, 10 - вторичная флоэма, 11 - вторичная кора, 12 - феллоген, 13 - феллема.
Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными организмами, в связи с тем что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем.
Большое значение для формирования корневых систем играет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватываются атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков.
Важное значение для развития корневых систем имеет аэрация . Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания . Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений – их усиленному ветвлению.
Строение корня растения
Основные функции корня: обеспечивает закрепление растения в почве, всасывание почвенного водного раствора солей и транспорт его к надземным частям растения.
Дополнительные функции: запасание питательных веществ, фотосинтез, дыхание, вегетативное размножение, выделение, симбиоз с микроорганизмами, грибами. Первые настоящие корни появились у папоротникообразных.
Зародыш корня называется зародышевым корнем и закладывается одновременно с почкой в зародыше семени.
У растений различают:
Главный корень. Он образуется из зародышевого и сохраняется на протяжении всей жизни. Всегда один.
Боковые корни. Ответвляются от корней (главного, дополнительных, боковых). Образуют при ветвлении корни 2-го, 3-го и т. д. порядка.
Придаточные корни . Образуются в любой части растения (стебле, листьях).
Совокупность всех корней растения образует корневую систему. Корневая система формируется в течение всей жизни растения. Ее формирование обеспечивают преимущественно боковые корни. Различают два типа корневой системы: стержневую и мочковатую.
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Меристемы- образовательные ткани- расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля).
Определение размеров корневых систем требует специальных методов. Оказалось, что общая поверхность корней обычно превышает поверхность надземных органов в 104-150 раз. При выращивании одиночного растения ржи было установлено, что общая длинна его корней достигает 600 км., при этом на них образуется 15 млрд. корневых волосков. Эти данные говорят об огромной потенциальной способности к росту корневых систем. Однако эта способность не всегда проявляется. При росте растений в фитоценозах, с достаточно большой густотой их строение, размеры корневых систем заметно уменьшаются.
С физиологической точки зрения корневая система не однородна. Довольно не вся поверхность корня участвует в поглощении волы. В каждом корне различают несколько зон (рис.1). Правда, не всегда все зоны выражены одинаково четко.
Окончание корня с наружи защищено корневым чехликом, напоминающим округлый колпачок, таящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика слущиваются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усилено делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона-зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки, затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а так же расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.
Рис. 1. Схема строения корня:
А - продольный разрез: 1-корневой чехлик; 2- меристема; 3-зона растяжения; 4- зона корневых волосков; 5- зона ветвления;
Б - поперечный разрез (по М.Ф. Даниловой): 1 - ризодерма; 2 - корневой волосок; 3 - паренхима; 4 - эндодерма; 5- пояски Каспари; 6 - перицикл; 7 - флоэма; 8 - ксилема. Пунктирные стрелки- пути передвижения веществ, поглощаемых из наружного раствора. Сплошные стрелки путь растворов по симпласту; прерывистые - путь по апопласту.
Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системных крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток- эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином. При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения, как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня.. При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление волосков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является основной зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет 5-10 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче.
Корень по длине можно разделить на несколько участков, имеющих различное строение и выполняющих различные функции. Эти участки называют зонами корня. Выделяют корневой чехлик и следующие зоны: деления, растяжения, всасывания и проведения.
Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы конуса нарастания. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует самый поверхностный слой с корневыми волосками - ризодерма (эпиблема). Защитную функцию выполняет ниже лежащая ткань - экзодерма.
В первичном строении корня выделяют три части: ризодерму, первичную кору и осевой (центральный) цилиндр.
На первичную кору приходится основная масса первичных тканей корня. Ее клетки накапливают крахмал и другие вещества. Эта ткань содержит многочисленные межклетники, имеющие значение для аэрации клеток корня. Наружные клетки первичной коры, лежащие непосредственно под ризодермой, называются экзодермой. Основная масса коры (мезодерма) образована паренхимными клетками. Самый внутренний слой носит название эндодермы. Это ряд плотно сомкнутых клеток (без межклетников).
Центральный или осевой цилиндр (стела) состоит из проводящих тканей, окруженных одним или несколькими слоями клеток - перициклом.
Внутренняя часть центрального цилиндра у большинства растений занимает сплошной тяж первичной ксилемы, дающий к перициклу выступы в виде ребер. Между ними размещаются тяжи первичной флоэмы.
У двудольных и голосеменных растений уже в раннем возрасте в центральном цилиндре корня между ксилемой и флоэмой появляется камбий, деятельность которого приводит к вторичным изменениям и в конечном итоге формируется вторичная структура корня. К центру камбий откладывает клетки вторичной ксилемы, а к периферии - клетки вторичной флоэмы. В результате деятельности камбия первичная флоэма оттесняется кнаружи, а первичная ксилема остается в центре корня.
Вслед за изменениями в центральном цилиндре корня происходят изменения в коровой части. Клетки перицикла начинают делиться по всей окружности, в результате чего возникает слой клеток вторичной меристемы - феллогена (пробкового камбия). Феллоген, в свою очередь, делясь, откладывает наружу феллему, а внутрь - феллодерму. Образуется перидерма, пробковый слой которой изолирует первичную кору от центрального цилиндра. В результате вся первичная кора отмирает и постепенно сбрасывается; наружным слоем корня становится перидерма. Клетки феллодермы и остатки перицикла в дальнейшем разрастаются и составляют паренхимную зону, которую называют вторичной корой корня (рис. 2).
При развитии запасающей паренхимы главного корня происходит формирование запасающих корней или корнеплодов. Различают корнеплоды:
1. Монокамбиальные (редька, морковь) - закладывается только один слой камбия, а запасные вещества могут накапливаться либо в паренхиме ксилемы (ксилемный тип - редька), либо в паренхиме флоэмы (флоэмный тип - морковь);
2. Поликамбиальные - через определенные промежутки времени происходит заложение нового слоя камбия (свекла).
Рис. 2. Переход от первичного строения корня к вторичному:
1 - первичная флоэма, 2 - первичная ксилема, 3 - камбий, 4 - перицикл, 5 - эндодерма, 6 - мезодерма, 7 - ризодерма, 8 - экзодерма, 9 - вторичная ксилема, 10 - вторичная флоэма, 11 - вторичная кора, 12 - феллоген, 13 - феллема.
Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными организмами, в связи с тем что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем.
Большое значение для формирования корневых систем играет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватываются атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков.
Важное значение для развития корневых систем имеет аэрация . Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания . Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений – их усиленному ветвлению.
Видоизменение корней
Корень растения выполняет множество функций. Наиболее важные из них для жизнедеятельности растения – удержание растения в почве и поглощение воды с растворенными минеральными веществами. Помимо вышеуказанных корням свойственно выполнение еще и ряда других функций, из-за чего происходит видоизменение их структуры. Вследствие метаморфоз видоизмененный корень теряют схожесть с обычным корнем.
Корнеплод
У некоторых двулетних растений (репы, петрушки, моркови) корень преобразуется в корнеплод, который представляет собой утолщенный придаточный корень. В формировании корнеплода принимают участие главный корень и нижний участок стебля. В структуре корнеплода главное место принадлежит запасающей основной ткани.
В зависимости от строения корня различают три типа корнеплодов: морковный, свекольный и редечный.
1. Корнеплоды морковного типа - овощи с удлиненной формой корня, который может быть цилиндрическим, коническим, удлиненно - коническим, веретенообразным и тупым или острым концом. У корнеплодов этого типа четко разграничены кора (флоэма) и сердцевина (ксилема). Между ними находится пробковый камбий. Сверху корнеплод покрыт естественной перидермой. По составу и количеству питательных веществ кора более ценная, чем сердцевина. К корнеплодам этого типа относится морковь, петрушка, сельдерей, пастернак.
2. Корнеплоды свекольного типа - овощи с округлыми, кругло-плоскими, овальными или удлиненными корнеплодами. Представлены столовой и сахарной свеклой. В качестве овощной культуры используется лишь столовая свекла. У корнеплода темно-красная мякоть с кольцами более светлого тога, что обусловлено чередованием тканей ксилемы (светлых колец) и флоэмы (темных колец). Чем меньше удельный вес занимают, такни ксилемы, тем выше пищевая ценность свеклы.
3.
Корнеплоды редечного типа
- овощи с округленными, реповидными, удлиненно-коническими корнеплодами. Особенностью их внутреннего строения является радиальное расположение вторичной ксилемы, флоэмы и паренхимной ткани. Камбиальный слой находится непосредственно под перидермой. К корнеплодам этого типа относятся редька, редис, брюква и репа.
Для корнеплодов всех типов характерны общие морфологические признаки: головка в верхней части с черешками листьев и почками в основании, корневое тело (основная съедобная часть) и кончик корня (основной), а у корнеплодов свекольного типа наличие боковых корешков. У остальных корнеплодов тонкие боковые корешки при уборке легко отрываются и, как правило, отсутствуют.
Особенность всех корнеплодов - их способность заживлять механические повреждениям путем суберинизации клеток, а так же их легкая усвояемость. Наиболее легко увядающими являются корнеплоды морковного типа, редис, наименее - свекла, редька, репа и брюква.
Корневые клубни (корневые шишки)
Многие покрытосеменные растения имеют на корнях корневые клубни (или корневые шишки). Их происхождение связано с утолщением придаточных и боковых корней. Корневые клубни – это вместилище питательных веществ. Их также используют для вегетативного размножения растений.
Самое известное из растений с типичным корнеклубнем - георгина. Вздутые корневые шишки отходят из одной точки - основания старого стебля. Эти видоизмененные корни обеспечивают растение питательными веществами. В период роста от них отрастают собственные тонкие корешки, которые добывают из почвы воду и питательные вещества. Размножают растения с корневыми шишками отдельными шишками с почкой (глазком) на конце (георгина, эремурус, кливия, лютик).
Корни-зацепки, ходульные корни, воздушные корни, досковидные корни, корни-присоски
Корни-зацепки
– своеобразные придаточные корни, позволяющие растению легко прикрепляться к любой опоре. Такие корни встречаются у плюща и других лазающих растений.
Удивительное явление в природе – ходульные корни, которые выполняют у растения роль опоры. Такие корни имеют способность противодействовать сильным нагрузкам, благодаря наличию механических тканей во всех отделах. Встречаются у растений рода пандаус, призрастающих на океанических островах в тропиках, где господствуют сильные ураганные ветры.
Воздушные (дыхательные) корни имеются у тропических деревьев, которые растут на почвах, обедненных кислородом, пресноводных тропических болотах. Это боковые корни, располагающиеся над землей. Благодаря воздушным корням, кислород и вода поглощаются растением из воздуха в условиях повышенной влажности. Так дыхательные корни обеспечивают функцию дополнительного дыхания.
Досковидные корни - это вертикальные надземные корни, характерные для крупных деревьев, растущих в дождевых тропических лесах. Развиваются у основания ствола, выглядят как доски, прилегающие к стволу, обеспечивая растению дополнительную опору.
Микориза
Микориза представляет собой симбиоз корней высших растений с гифами грибов. Это сожительство выгодно для обоих организмов, так как гриб получает от растения готовые органические вещества, а растение всасывает от гриба воду с минеральными веществами. Микориза часто встречается на корнях высших растений, особенно древесных. Можно представить грибные гифы, переплетающиеся с толстыми корнями деревьев, как корневые волоски, поскольку функции их аналогичны.
Большая часть многолетних растений имеет микоризу. Предполагается, что микориза является одним из факторов, способствовавших прогрессу цветковых растений. Растения, питающиеся с помощью микоризных грибов, называются микотрофными.
Различают два основных типа микоризы: эктотрофную и эндотрофную
. Гифы эктотрофной микоризы охватывают корень только снаружи, иногда проникая в межклетники коровой паренхимы. Эктомикориза обычная у многих древесных (сосна, берёза, дуб, бук и др.), а также ряда травянистых сельскохозяйственных растений, особенно у злаков. Гриб поселяется на проросшем корешке семени и в дальнейшем развитии, особенно в фазе кущения, обеспечивает обильное почвенное питание растения.
Эндотрофная микориза встречается более широко. Она характерна для большинства цветковых. Эндомикориза не образует грибной чехол вокруг корня, корневые волоски не отмирают, но гифы проникают в корень и внедряются в клетки коровой паренхимы.
У эпифитных тропических орхидей и некоторых других растений существуют так называемые воздушные корни. Они покрывают многослойной эпиблемой, называемой веламеном. Веламен выполняет иногда фотосинтезирующую функцию, а позже может участвовать в поглощении атмосферной влаги, образуя мёртвый губчатый гигроскопичный покров корня.
Без микоризы мицелий симбиотических грибов может определенное время существовать в почве, но при этом никогда не будет образовывать плодовых тел. Поэтому в искусственных условиях нельзя получить плодовые тела белого гриба, сыроежковых грибов, мухоморов - все они являются микоризообразователями, и без определенной древесной породы не будут плодоносить. В свою очередь растение без своих грибных симбионтов развивается плохо, медленно, легко подвергается заболеваниям и даже может погибнуть.
Огромное значение микориза имеет в тропических дождевых лесах. Из-за сильного промывного режима (ежедневное выпадение осадков) эти леса практически лишены почвы (все питательные вещества из почвы вымываются). Перед растениями остро встает проблема питания. В то же время, свежей органики очень много: опавшие ветви, листья, плоды, семена. Но эта органика недоступна высшим растениям, и они вступают в тесный контакт с сапротрофными грибами. Таким образом, основным источником минеральных веществ в этих условиях является не почва, а почвенные грибы. Минеральные вещества поступают в корень непосредственно из гиф микоризных грибов, именно поэтому для растений дождевого леса характерна поверхностная корневая система. О том, насколько эффективно работает микориза, можно судить хотя бы по тому, что тропические дождевые леса являются самыми продуктивными на Земле сообществами, здесь развивается максимально возможная биомасса.
Бактериальные клубеньки
Также растения могут взаимовыгодно сожительствовать с азотфиксирующими бактериями. Так на корнях высших растений появляются бактериальные клубеньки – видоизмененные боковые корни, которые имеют приспособления для симбиоза с бактериями. Через корневые волоски бактерии попадают внутрь молодых корней и провоцируют формирование клубеньков. Роль данных бактерий состоит в том, что они преобразуют азот из воздуха в минеральную форму таким образом, что он становится доступным для усвоения растениями. Растения выполняют функцию защиты для бактерий от конкурирующих видов почвенных бактерий. Также бактерии питаются веществами из корней высших растений. Зафиксировано появление клубеньковых бактерий, в основном, на корнях растений из семейства бобовые. Поэтому семена бобовых обогащены белком, а в сельском хозяйстве представителей этого семейства используют в севообороте, чтобы обогатить азотом почву.
Во влажных тропических лесах имеются растения, произрастающие в кронах деревьев. Их называют эпифитами (от греч. «эпи» - на и «фитон» - растение). Корни таких растений называют воздушными, так как они обычно свешиваются с ветвей и способны подобно промокательной бумаге поглощать влагу и мелкие частицы, находящиеся в воздухе. К эпифитам относят, например, многие орхидеи


