Физиология мышц их свойства строение функции. Реферат Механизм сокращения скелетной мышцы. Роль ионов кальция. Расслабление скелетной мышцы. Значение АТФ
Мышечная ткань выполняет следующие функции:
Обеспечение двигательной активности – целенаправленное поведение есть наиболее эффективная форма приспособления.
Обеспечение особых, присущих только человеку функций – прежде всего это коммуникативная функция , выражающаяся в виде устной и письменной речи.
Выполнение дыхательной функции – экскурсия грудной клетки и диафрагмы.
Участие в процессах теплообразования – терморегуляторный тонус, мышечная дрожь.
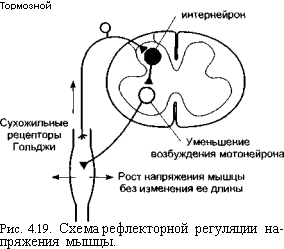
Специфическим свойством всех типов мышц является сократимость – способность сокращаться, то есть укорачиваться или развивать напряжение. Для реализации этой способности мышца использует два своих дополнительных свойства – возбудимость и проводимость .
Скелетные мышцы называют также произвольными , так как их сокращением можно управлять по собственному желанию. Они полностью лишены автоматизма и не способны работать без управляющей импульсации из ЦНС. Гладкие мышцы по собственному желанию не сокращаются, поэтому их называют также непроизвольными .
Морфофункциональная характеристика скелетной мышцы . Скелетная мышца состоит из многоядерных мышечных волокон. Толщина волокна составляет от 10 до 100 мкм. Длина волокон колеблется от нескольких мм до нескольких сантиметров.
Количество мышечных волокон становится постоянным на 4-5 месяце постнатального развития. В последующем увеличивается лишь диаметр и длина волокон (например, под действием тренировок – функциональная гипертрофия).
Мышечное волокно покрыто сарколеммой. В саркоплазме мышечного волокна имеются следующие внутриклеточные элементы: ядра, митохондрии, белки, капельки жира, гранулы гликогена, фосфатсодержащие вещества, различные малые молекулы и электролиты. От поверхности сарколеммы внутрь волокна отходят Т-трубочки, которые обеспечивают её взаимодействие с саркопламатическим ретикулумом. Саркоплазматический ретикулум представляет собой систему связанных между собой цистерн и отходящих от них в продольном направлении канальцев , расположенных между миофибриллами. Крайние цистерны ретикулума связаны с Т-трубочками. В цистернах содержатся ионы кальция, необходимые для осуществления процесса сокращения. Внутри мышечного волокна тянется масса нитей – миофибрилл, которые являются частью механизма процесса сокращения. Каждая миофибрилла состоит их протофибрилл, которые расположены параллельно друг другу и имеют белковую природу.
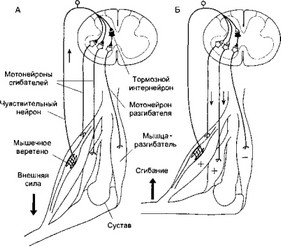
Различают два вида внутримышечных нитей: тонкие актиновые и толстые миозиновые . Актиновые нити состоят из двух субъединиц - скрученных в виде спирали волокон, каждое их которых образовано соединенными молекулами глобулярного белка актина. Кроме актина в состав тонких нитей входят регуляторные белки тропомиозин и тропонин . Эти белки в невозбужденной мышце препятствуют связи актина и миозина, поэтому мышца в покое находится в расслабленном состоянии.
Рис.1. Схема пространственной организации сократительных и регуляторных белков в исчерченной мышце.
Каждая миозиновая нить окружена шестью актиновыми нитями. Эти нити образуют своего рода цилиндр, внутри которого располагается миозиновая нить. Поперечные мостики миозиновой нити направлены в разные стороны, поэтому они взаимодействуют со всеми актиновыми протофибриллами. В свою очередь, каждая нить актина контактирует с тремя миозиновыми филаментами.
Миофибриллы включают в себя последовательно соединенные блоки – саркомеры , отделенные друг от друга Z-полосками. Саркомер является сократительной единицей мышечного волокна. Миофибриллы отдельного мышечного волокна связаны таким образом, что расположение саркомеров совпадает, это визульно делает мышцу поперечнополосатой. Миозиновые протофибриллы образуют наиболее темную часть саркомера – А-диск (анизотропный – сильно поляризует видимый свет). Более светлый участок в центре А-диска называют Н-зоной. Светлый участок саркомера между двумя А-дисками называют I-диском (изотропный – не поляризует свет). Он образован актиновыми протофибриллами, идущими в обе стороны от Z-полосок.
Таким образом, каждый саркомер имеет два набора тонких нитей, прикрепленных к Z-полоскам, и один комплект толстых нитей, сосредоточенных в А-диске. В расслабленной мышце концы толстых и тонких филаментов в разной степени перекрывают друг друга на границе между А- и I-дисками. В световом микроскопе зона перекрытия в А-диске представляется темнее, чем центральная Н-зона, лишенная актиновых нитей.
По морфологическим признакам и физиологическим свойствам различают быстрые и медленные волокна.
Быстрые (белые ) мышечные волокна содержат больше миофибрилл, в них мало митохондрий, миоглобина и жиров. Капиллярная сеть у них редкая. Для них характерна высокая скорость сокращения. Они обладают малой выносливостью.
Медленные (красные ) мышечные волокна имеют меньший диаметр. В цитоплазме много митохондрий, миоглобина и жирных кислот. Эти волокна окружены густой сетью капилляров. Скорость сокращения меньше, чем у белых волокон. Волокна этого типа выполняют в течение длительного периода времени, утомление в них развивается медленнее . Они выполняют функцию поддержания тонуса.
Имеются также истинные тонические мышечные волокна, которых локализуются по 7-10 синапсов, принадлежащие нескольким мотонейронам. ПД в этих волокнах потенциалы концевой пластинки не формируют, а запускают мышечное сокращение непосредственно. Тонических волокон в организме человека мало, они имеются только в глазодвигательных мышцах и мышцах среднего уха.
Механизм сокращения скелетной мышцы
. Укорочение мышцы является результатом сокращения множества саркомеров, которые последовательно соединены в миофибриллах. При укорочении мышцы актиновые нити скользят относительно миозиновых, в результате чего длина саркомера уменьшается, размер А-дисков остается постоянным, I-диски и Н-зоны уменьшаются. Длина самих же нитей остается неизменной. Совокупность процессов, обеспечивающих связь между возбуждением и сокращением, называется элекромеханическим сопряжением
. Электромеханическое сопряжение состоит из следующего ряда процессов:
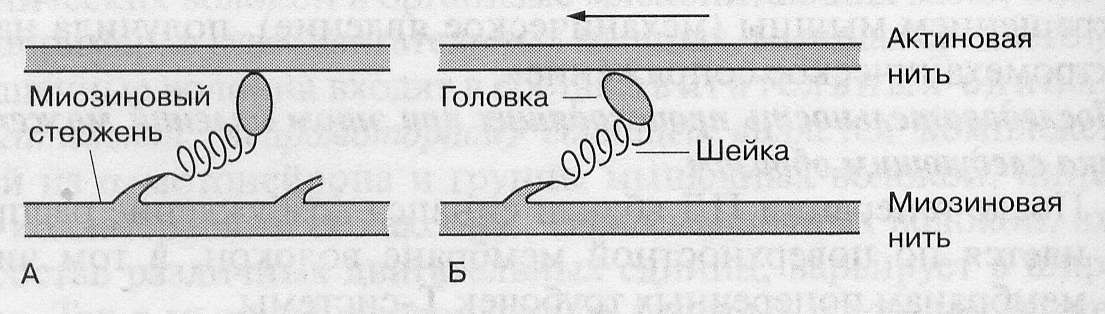
Движение сократительных белков происходит не рывками, а равномерно и плавно, так как в каждый момент какое-то количество миозиновых головок взаимодействует с активными участками актиновых нитей , другие подвергаются конформации, а третьи находятся на стадии восстановления исходной пространственной ориентации.
При расслаблении мышцы, описанные выше процессы, протекают в обратной последовательности: реполяризация сарколеммы и Т-трубочек вызывает закрытие кальциевых каналов мембраны саркоплазматического ретикулума и включение её кальциевых насосов, которые возвращают ионы кальция в цистерны ретикулума. Снижение концентрации ионов кальция в межфибриллярном пространстве вызывает обратную конформацию тропонина, в результате чего тропомиозиновые нити изолируют активные участки актиновых нитей. Контакт миозиновых головок с актином разрывается. Актиновые нити под действием веса мышцы и тяги эластических элементов скользят в обратном направлении и размер саркомера восстанавливается.
Энергетическое обеспечение мышечного сокращения . На функционирование мускулатуры в состоянии максимального физиологического покоя организм тратит около 20% всех своих энергоресурсов. При мышечной работе эти энергозатраты многократно возрастают. Источником энергии для жизнедеятельности мышечной ткани является АТФ.
Энергия АТФ расходуется для осуществления следующих процессов:
Работа натрий-калиевого насоса, который обеспечивает поддержание градиентов концентрации ионов калия и натрия по разные стороны клеточной мембраны; формирование за счет этого потенциала покоя и сохранение свойств возбудимости и проводимости.
Взаимодействие актиновых и миозиновых нитей при сокращении мышцы. При этом энергия АТФ расходуется, во-первых, на процесс скольжения, во-вторых, на весь гребковый цикл: присоединения АТФ к поперечному мостику миозиновой нити происходят в конце предыдущего гребка, что обеспечивает «отрыв» головки от актина, возможность прикрепления её к новому участку актиновой нити, и собственно гребок. При отсутствии АТФ головки миозина не могут оторваться от актина – мышца напряжена. Таков, в частности, механизм трупного окоченения.
Работа кальциевого насоса, активация которого вызывает понижение концентрации ионов кальция в саркоплазме, что приводит к расслаблению мышечного волокна.
Фосфогенная энергетическая система обеспечивает ресинтез АТФ за счет энергии, освобождающейся при распаде имеющегося в мышцах креатинфосфата с образованием креатина: АДФ + КФ →АТФ + К. Эотот путь ресинтеза самый быстрый, но обладает небольшой емкостью. Работа до 5 секунд.
Анаэробная гликолитическая энергетическая система осуществляет ресинтез АТФ за счет энергии анаэробного расщепления глюкозы до молочной кислоты. Эото способ является быстрым и эффетивным, но емкость его не достаточно большая. Работа несколько десятков секунд.
Аэробные энергетические системы обеспечивают ресинтез АТФ в митохондриях мышечных волокон. В качестве субстратов используются углеводы и жирные кислоты. Обеспечивает работу мышцы в течение нескольких часов. Он более медленный и менее мощный, но является основным способом энергетического обеспечения работы мышц в реальных условиях.
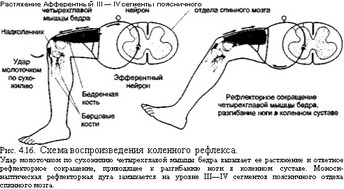
Одиночное сокращение мышцы – это сокращение, которое возникает при действии на мышцу одиночного электрического или нервного импульса. На кривой одиночного сокращения выделяют три основных фазы: 1. латентный период – время нанесения раздражения до начала сокращения. В этот период возникает возбуждение мышечных волокон и его распространение вдоль мембраны. Продолжительность варьирует до 10 мс; 2. фаза укорочения (или развитие напряжения) – около 50мс; 3. фаза расслабления - около 60 мс.
Тетаническое сокращение – это длительное укорочение мышцы, возникающее под действием ритмического раздражения. В его основе лежит суммация одиночных сокращений. При тетаническом сокращении амплитуда больше, чем при одиночном сокращении, так как повторные потенциалы действия возникают прежде, чем саркоплазматический ретикулум сможет удалить ранее высвобожденный кальций, поэтому уровень последнего в гиалоплазме повышается , активное состояние продлевается, увеличивается количество работающих мостиков и, как результат, усиливается сила сокращения. Для возникновения тетануса необходимо, чтобы интервал между стимулами был больше рефрактерного периода, но короче всей длительности сократительного ответа. Тетаническое сокращения имеет два вида: зубчатый тетанус и гладкий. Если повторить раздражение в фазу расслабления, то получится зубчатый тетанус , если же в фазу укорочения – то гладкий . При некоторой достаточно высокой частоте раздражения нерва амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус называется оптимумом . Для развития оптимума необходимо, чтобы повторные раздражители поступали к мышце после завершения периода рефрактерности, вызванного предыдущим раздражителем. Если повышать дальше частоту раздражения, то наступает состояние, которое называется пессимумом Введенского – формируется блок проведения возбуждения в нервно-мышечном синапсе и мышца вместо того, чтобы продолжать возбуждаться, расслабляется, сколько бы мы её не раздражали. В естественных условиях ввиду асинхронности работы мотонейронов сокращение мышцы напоминает гладкий тетанус.
Изотонический режим – это сокращение мышцы , при котором её волокна укорачиваются, но напряжение остается постоянным. В реальных условиях чисто изотоническое сокращение отсутствует.
Изометрический режим – сокращение мышцы, при котором её длина не изменяется, но развиваемое ею напряжение возрастает. Например, поднятие груза, который больше силы мышцы.
Ауксотонический режим – это сокращение мышцы, при котором одновременно изменяется длина и напряжение. Этот режим характерен для натуральных двигательных актов.
Δ Сила мышцы или общая сила мышцы , определяется максимальным напряжением в ньютонах, которое она может развить. Удельная сила мышцы – отношение общей силы в ньютонах к физиологическому поперечному сечению мышцы (Н/см 2). С помощью динамометров можно измерить «кистевую», «становую» силу, силу сгибателей и т.д. Сила мышцы зависит от
Анатомического строения – если волокна мышцы расположены под углом, то её сила больше (перистые мышцы).
Объема растяжения – при умеренном растяжении сократительный эффект мышцы увеличивается, при сильном – уменьшается.
Силы стимула : одиночное мышечное волокно реагирует на раздражение по правилу «все или ничего». Сокращение целой мышцы зависит от силы стимуляции до тех пор, пока не будут вовлечены в процесс все волокна мышцы . Затем мышца перестает выдавать увеличение амплитуды сокращения.
Удельная сила мышцы зависит от входящих в её состав белых и красных волокон . Чем больше быстрых (белых) волокон, тем выше удельная сила.
Δ Мощность мышцы – работа, совершаемая в единицу времени. Выражается в ваттах – Дж/с 2 . Максимальная мощность соответствует выполнению наибольшего объема работы в течение минимального отрезка времени, но в этом случае быстро развивается утомление. Поэтому необходимо использовать правило средних нагрузок.
Физиология гладкой мышцы . Морфофункциональные особенности гладких мышц следующие:
Сократительные белки (актин и миозин) гладких мышц ориентированы не столь упорядоченно, Z-мембраны и саркомеры в них отсутствуют. В клетке имеется одно ядро, митохондрий мало. Сакркоплазматический ретикулум состоит из плоских везикул .
Синапсы между аксонами вегетативных эфферентных нейронов и гладкомышечными клетками организованы иначе, чем соматические – нервно-мышечные. Терминали аксонов ганглионарных нейронов образуют утолщения, из которых выделяется медиатор. Этот медиатор распределяется по межклеточному пространству и контактирует с мембранами нескольких гладкомышечных клеток. Причем, постсинаптические рецепторы располагаются равномерно по всей поверхности данных клеток. Такие синапсы наиболее характерны для симпатической нервной системы.
Возбудимость – потенциал покоя и потенциал действия большинства гладкомышечных клеток имеет более высокие значения и более значительную продолжительность соответственно, чем миоциты скелетной мускулатуры.
Изолированные отдельные гладкие волокна не возбуждаются и не сокращаются. Взаимодействие между отдельными клетками осуществляется благодаря щелевидным контактам, которые обладают низким сопротивлением.
У гладкой мускулатуры имеются структурно-функциональные единицы, которые состоят из группы клеток, объединенных нексусами и электрическими полями. Структурно-функциональная единица гладкой мышцы сокращается как единое целое. Активность АТФазы гладкого миоцита меньше, чем активность АТФазы скелетного миоцита. Поэтому гладкая мышца сокращается медленнее, чем скелетная, практически не утомляется и меньше расходует АТФ.
Гладким мышцам присуща автоматия. Например, пейсмекерам. В основе автоматии лежит спонтанно возникающая медленная поляризация. Спонтанная деполяризации обусловлена диффузией в клетку кальция .
Гладкая мускулатура обладает пластичностью. Пластичность способствует растяжению, которое, в свою очередь, способствует возникновению спонтанного потенциала действия. Например, растяжение мочевого пузыря ведет к его сокращению и эвакуации мочи.
Электромеханическое сопряжение гладких мышц также имеет свои особенности. Во-первых, запасы кальция в гладкомышечном волокне незначительны. Поэтому инициация сокращения осуществляется в основном за счет внеклеточного кальция. Для того чтобы кальция хватило на запуск процесса возбуждения природа увеличила проницаемость мембраны гладкомышечного миоцита для кальция при формировании ПД. Во-вторых, тропонина в глаких мышцах нет , а есть белок кальмодулин, который связывается с кальцием. Комплекс кальмодулин-кальций активирует фермент, который переносит фосфатную группу с АТФ на головку мостика миозина. И уже фосфолирированная головка миозина взаимодействует с актином. В-третьих, расслабление гладкой мышцы обусловлено инактивацией кальциевых каналов. Из гиалоплазмы начинает уходить кальций и миозиновые головки заканчивают своё взаимодействие с актином.
Ресинтез молекул АТФ в гладких мышцах идет в основном по пути анаэробного гликолиза.
В гладкой мышце , как и в скелетной, сократительный процесс активируется ионами кальция, однако источники этих ионов различны. Различие заключается в том, что саркоплазматический ретикулум, обеспечивающий практически все количество ионов кальция для сокращения скелетной мышцы, в большинстве гладких мышц очень слабо развит.
Вместо этого почти все ионы кальция , вызывающие сокращение, входят в мышечную клетку из внеклеточной жидкости во время потенциала действия или под влиянием другого стимула.
Поскольку концентрация ионов кальция во внеклеточной жидкости выше 10 М, а внутри гладкомышечной клетки - менее 10 М, открытие кальциевых каналов вызывает быструю диффузию ионов кальция в клетку. Время, необходимое для осуществления этой диффузии, в среднем составляет 200-300 мсек. Его называют латентным периодом процесса сокращения. Для гладкой мышцы латентный период примерно в 50 раз больше, чем для скелетной мышцы.
Роль саркоплазматического ретикулума гладкомышечных волокон . На рисунке показаны немногочисленные слаборазвитые трубочки саркоплазматического ретикулума, которые лежат около клеточной мембраны в некоторых более крупных гладкомышечных клетках. К этим трубочкам примыкают небольшие инвагинации клеточной мембраны, называемые кавеолами. Кавеолы представляют собой рудиментарный аналог системы поперечных трубочек скелетной мышцы.
Появление потенциала действия в мембране кавеол , как полагают, ведет к выделению ионов кальция из примыкающих саркоплазматических трубочек так же, как в скелетных мышцах потенциалы действия в области Т-трубочек вызывают выделение ионов кальция из цистерн саркоплазматического ретикулума. В целом, чем больше развит сарко-плазматический ретикулум в волокне гладкой мышцы, тем быстрее оно сокращается.
Влияние на сокращение гладких мышц изменений внеклеточной концентрации ионов кальция. Изменение концентрации ионов кальция во внеклеточной жидкости практически не влияет на силу сокращения скелетных мышц, однако для большинства гладких мышц это не так. При снижении концентрации кальция во внеклеточной жидкости примерно до 1/3-1/10 нормы сокращения гладких мышц обычно прекращаются. Следовательно, сила сокращения гладкой мышцы в высокой степени зависит от концентрации ионов кальция во внеклеточной жидкости.
Для расслабления гладкой мышцы необходим кальциевый насос. Для расслабления гладкой мышцы после ее сокращения необходимо удалить ионы кальция из внутриклеточной жидкости. Это удаление осуществляется с помощью кальциевого насоса, который выкачивает ионы кальция из гладкомышечного волокна назад во внеклеточной жидкости или саркоплазматический ретикулум (при его наличии). По сравнению с быстродействующим насосом саркоплазматического ретикулума скелетной мышцы кальциевый насос гладкомышечного волокна действует медленно.
В связи с этим одиночное сокращение гладкой мышцы часто продолжается в течение секунд, а не сотые или десятые доли секунды, что характерно для скелетной мышцы.
Вернуться в оглавление раздела " "
Скелетная мышца представляет собой комплекс отдельных многоядерных мышечных клеток или волокон, которые состоят из функциональных единиц - миофибрилл, осуществляющих механизм сокращения в результате взаимодействия белковых филаментов миозина и актина. Основной характеристикой скелетной мышечной ткани является ее способность под контролем со стороны нервной системы развивать силу сокращения и укорочение. Степень механического напряжения и укорочения мышцы обусловлена активацией числа двигательных единиц, образующих целую мышцу. Двигательной единицей скелетной мышцы называется количество двигательных (моторных) мышечных волокон, которые иннервируются одним двигательным нейроном спинного мозга.
Скелетное мышечное волокно представляет собой вытянутую многоядерную клетку, которая содержит сократительные элементы, или миофибриллы. В каждом мышечном волокне содержится до 1000 миофиб- рилл и более, толщиной 1 - ![]() alt="" />3 мкм. Отдельная миофиб- рилла состоит, в свою очередь, из множества толстых и тонких миофиламентов - нитей белка миозина и белка актина соответственно.
alt="" />3 мкм. Отдельная миофиб- рилла состоит, в свою очередь, из множества толстых и тонких миофиламентов - нитей белка миозина и белка актина соответственно. ![]()
 Взаимодействие между толстыми и тонкими миофила- ментами является физической основой мышечного сокращения. Миофиламен- ты окружены цитоплазмой, называемой саркоплазмой.
Взаимодействие между толстыми и тонкими миофила- ментами является физической основой мышечного сокращения. Миофиламен- ты окружены цитоплазмой, называемой саркоплазмой.
Кроме того, в состав нитей актина входят белки - тро- понин и тропомиозин, которые необходимы для развития процессов сокращения и расслабления мышцы. Каждая миофибрилла содержит примерно 1500 миози- новых и около 3000 актино- вых миофиламентов.
По всей длине мышечной клетки миофиламенты миофибриллы разделены на структурно-функциональные единицы, которые называются саркомерами (рис. 2.25). Саркомеры отделены друг от друга Z-мем-
бранами, к которым прикреплены тонкие актиновые филаменты. Расположены толстые и тонкие филаменты в саркомере гексамодально, т. е. вокруг каждого толстого миозинового филамента расположено шесть тонких ак- тивновых филаментов.
Нити миозина лежат в центре саркомера и выглядят в световом микроскопе как темная полоска, которая называется анизотропным А-диском. По обе стороны от A-диска лежат тонкие нити актина, которые в световом микроскопе образуют светлые изотропные диски. Нити актина, которые образуют в саркомере два изотропных диска, тянутся до Z-мембран.
Функции миофиламентов
Сократительная способность скелетной мышцы обусловлена функцией мышечных белков актина и миозина, взаимодействие которых между собой является основой механизма сокращения скелетной мышцы.
Актиновые филаменты представляют собой двойные белковые спирали F-актина, каждая из которых образована примерно 200 единицами (мономерами) глобулярного G-актина. Каждый G-актин состоит из четырех субдоменов. Два больших субдомена (3-й и 4-й) актина выполняют функцию связывания между собой белковой спирали F-актина, а два маленьких (1-й и 2-й), локализованные на периферии глобулярного G-актина, являются местами взаимодействия каждого мономера G-актина с толстым филамен- 
 том. Основную роль в связывании актина с миозином во время сокращения мышцы выполняет субдомен 1, содержащий NH2 СООН терминали актина. Этот субдомен актина выполняет функцию связывающего центра. Поэтому каждый глобулярный G-актин представляет собой место связывания с миозином при сокращении мышцы. В состоянии покоя скелетной мышцы центры связывания на каждом глобулярном G-актине закрыты белками тропонином и тропо- миозином. Эти белки функционально связаны между собой и образуют тропонин-тропомиози- новый комплекс, который расположен вдоль всей двойной белковой спирали F-актина (рис. 2.26).
том. Основную роль в связывании актина с миозином во время сокращения мышцы выполняет субдомен 1, содержащий NH2 СООН терминали актина. Этот субдомен актина выполняет функцию связывающего центра. Поэтому каждый глобулярный G-актин представляет собой место связывания с миозином при сокращении мышцы. В состоянии покоя скелетной мышцы центры связывания на каждом глобулярном G-актине закрыты белками тропонином и тропо- миозином. Эти белки функционально связаны между собой и образуют тропонин-тропомиози- новый комплекс, который расположен вдоль всей двойной белковой спирали F-актина (рис. 2.26).
Тропомиозин представляет собой молекулу длиной 42 нм, образованную двумя белковыми?- цепями по типу спирали. Тропомиозин связан электростатически с актиновым филаментом. Однако элетростатическая связь между актиновым филаментом и тропо- миозином является плотной только во время расслабления скелетной мышцы. В присутствии ионов Са2+ эта связь становится менее плотной, тропомиозин становится подвижным и способен открывать места связывания на актиновом филаменте для поперечных мостиков миозина, что происходит во время сокращения.
В разных типах мышечной
ткани тропомиозин имеет различную подвижность относительно актина. Например, в скелетной мышце тропомиозин более подвижен, чем в сердечной мышце, что обусловливает отличие в сократительных свойствах этих двух типов поперечно-полосатых мышечных тканей. Подвижность тропо- миозина относительно актинового филамента регулируется тропонином.
Тропонин состоит из трех белковых субъединиц: С, I и Т. Субъединица С тропонина является своеобразным сенсором для ионов Са2+ в регуляции сокращения как в скелетной, так и в сердечной мышцах. Тропонин С имеет две глобулярные области - NH2 и СООН терминали. В скелетном мышечном волокне каждая из этих концевых областей тропонина С содержит два возможных места связывания ионов Са2+. Функция субъединицы I тропонина заключается в связывании тропонина с актином. Субъединица Т тропонина прочно удерживает вместе тропонин-тропомиозиновый комплекс на нитях актина. />В присутствии ионов Са2+ связь тропонина Т и тропомиозина относительно тонкой нити актина становится подвижной. Это приводит к открытию мест связывания на актиновом филаменте, с которым способны связываться поперечные мостики толстого филамента с началом процесса мышечного сокращения.
Толстый филамент представляет собой биполярный полимер белка миозина, который способен взаимодействовать с актином и развивать в результате этого взаимодействия силу и укорочение мышцы.
Двигательный белок миозин состоит из двух тяжелых и четырех легких полипептидных цепей. Тяжелые полипептидные цепи образуют спираль миозина. Глобулярные концевые NH2 области называются головками, с помощью которых происходит взаимодействие между центрами связывания на актиновом филаменте с миозином во время сокращения мышцы. Одна пара легких полипептидных цепей связана с каждой головкой. Основа толстого филамента представлена миозиновым стержнем, который скручен из множества белковых нитей. Образование миозинового стержня происходит в период развития мышечной ткани, благодаря функции белка С саркоме- ра, который является антителом для белка миозина.
В скелетных мышцах от миозинового стержня по его окружности отходят с равным интервалом в 14,3 нм пары головок поперечных, а расстояние между головками по одной линии толстого филамента составляет 43 нм. При этом каждая пара головок на филаменте миозина ориентирована на 180° относительно другой. Подобная «геометрия» расположения поперечных мостиков на толстом филаменте обусловливает их взаимодействие с любой из шести окружающих его тонких филаментов с интервалом в 14,3 нм.
Важным внутриклеточным элементом эластичности мышечных волокон является высокоэластичный белок титин, который формирует своеобразную сеть вокруг нитей актина и миозина в пределах каждого саркомера и придает ему механические свойства как в покое, так и при мышечном сокращении. Благодаря наличию белка титина в саркомере, в состоянии покоя сократительные элементы развивают напряжение при растяжении мышцы. Механизм сокращения скелетной мышцы
Во время укорочения мышцы уменьшается длина саркомеров. При этом филаменты актина и миозина, входящие в состав саркомеров, не изменяют своей длины. Уменьшение длины соркомеров происходит в результате движения (скольжения) тонких филаментов с двух сторон саркомера к его
центру относительно толстых мио- зи новых филаментов (рис. 2.27). Молекулы актина и миозина способны взаимодействовать между собой как в условиях in vivo, так и in vitro. В мышце эта связь миози- новых головок с актиновым фила- ментом является циклической и представляет собой многочисленные укорочения миозиновых поперечных мостиков, что поддерживает постоянство сокращения в целой активной мышце. 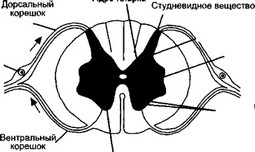
 Цикл поперечного мостика во время сокращения мышцы - это связывание, укорочение, высвобождение и вновь связывание поперечного мостика миозина с актиновым филаментом, что весьма напоминает собой человека, который на руках поднимается вверх по канату. В саркомере каждый миозиновый филамент находится в окружении шести актиновых филаментов. Миозиновые фила- менты вращаются во время взаимодействия с окружающими их шестью филаментами актина, а их поперечные мостики связываются лишь с тремя случайными тонкими миофиламентами. Согласно теории скольжения филаментов мышечного сокращения поперечные мостики миозина взаимодействуют с актином асинхронно, т. е. одни находятся «в захвате», в то время как другие «изменяют свой захват». Миозиновые головки, которые образуют поперечные мостики, расположены только на концах миозиновых филаментов. Поэтому при максимальном увеличении длины саркомеров, несмотря на раздражение мышцы, полностью отсутствует сокращение мышцы.
Цикл поперечного мостика во время сокращения мышцы - это связывание, укорочение, высвобождение и вновь связывание поперечного мостика миозина с актиновым филаментом, что весьма напоминает собой человека, который на руках поднимается вверх по канату. В саркомере каждый миозиновый филамент находится в окружении шести актиновых филаментов. Миозиновые фила- менты вращаются во время взаимодействия с окружающими их шестью филаментами актина, а их поперечные мостики связываются лишь с тремя случайными тонкими миофиламентами. Согласно теории скольжения филаментов мышечного сокращения поперечные мостики миозина взаимодействуют с актином асинхронно, т. е. одни находятся «в захвате», в то время как другие «изменяют свой захват». Миозиновые головки, которые образуют поперечные мостики, расположены только на концах миозиновых филаментов. Поэтому при максимальном увеличении длины саркомеров, несмотря на раздражение мышцы, полностью отсутствует сокращение мышцы.
Активность актомиозиновых мостиков, с помощью которых актиновые филаменты прикрепляются к миозину, представляет собой процесс, в ходе которого химическая энергия АТФ превращается в сокращение мышцы. Каждый отдельный поперечный мостик имеет структурные особенности, которые вовлечены в сокращение мышцы. Эти особенности включают двойную головку миозина, которая содержит АТФазную активность поперечного мостика, и способность связываться плотно с актином, и в то же время вращается наподобие шарнира относительно стержня поперечного мостика. В свою очередь стержень поперечного мостика также наподобие шарнира связан с миозиновым стержнем и обладает эластическими свойствами, заключающими в себе энергию, необходимую при развитии напряжения во время вращении головки поперечного мостика.
Поперечные мостики отходят от миозинового стержня таким образом, что образуют спираль вдоль толстного филамента с интервалом 14,3 нм
 00000000000
00000000000 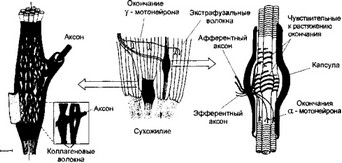

Актин
АТФ
Рис. 2.28. Цикл «замыкание-сокращение-размыкание» поперечного мостика миозина.
Стадия 1 - в саркоплазме низкая концентрация ионов кальция (менее 10~7 М), места связывания на филаменте актина блокированы тропонин-тропомиозиновым комплексом. Стадия 2 - коцентрация ионов кальция в саркоплазме возрастает до порогового уровня, поперечный мостик миозина связывается с актином. Связывание ускоряет гидролиз АТФ примерно в 200 раз с образованием АДФ и фосфата (Ф). Стадия 3 - энергия гидролиза АТФ вызывает движение головки поперечного мостика и одновременно растягивается эластичный компонент поперечного мостика. Стадия 4 - эластический компонент поперечного мостика укорачивается, что вызывает перемещение актинового филамента - это является «шагом» укорочения сарко- мера. Стадия 5 - с головкой поперечного мостика связывается новая молекула АТФ и поперечный мостик миозина высвобождается от связи с атином. При высокой концентрации ионов кальция в саркомплазме цикл «замыкание-сокращение-размыкание» будет неоднократно повторяться, а при удалении ионов кальция в саркоплазматический ретикулум скорость гидролиза АТФ резко снижается и тормозится прочное актомиозиновое взаимодействие.
между мостиками. Этот интервал столь мал, что каждой мостик должен многократно взаимодействовать с актином по мере продвижения вдоль актинового филамента, чтобы произошло существенное укорочение длины всей мышцы. />В состоянии покоя поперечный мостик расположен параллельно мио- зиновому стержню (рис.2.28), а его головка - перпедикулярно относительно последнего, и не прикреплена к актиновому филаменту (стадия 1). После того, как ионы Са2+ высвобождаются из саркоплазматического ретикулума (описано ниже) и активируют сокращение мышцы, активновый фи- ламент может связываться с миозиновой головкой (стадия 2). После связывания головка наклоняется (стадия 3), используя энергию, заключенную в миозин-АТФазном комплексе головки. Это является основной силой поперечного мостика, которая растягивает его эластический компонент для того, чтобы произвести энергию движения вдоль активного филамента (стадия 4). В результате головка мостика совершает движение на 40° к центру саркомера и перемещает при этом на 10 нм нить актина. В это время от головки высвобождаются АДФ и монофосфат, что необходимо для захвата головкой миозина новой молекулы АТФ. Только после этого головка
миозина отделяется от молекулы актина, необходимой для размыкания поперечного мостика с актином (положение 5).
Цикл будет продолжаться до тех пор, пока концентрация ионов Са++ в межфибриллярном пространстве не снижается до подпороговой величины, т. е. 1СН М, что приведет к расслаблению мышцы, или пока не будут исчерпаны запасы АТФ. В последнем случае в мышце возникает спазм или мышечная контрактура (rigor), при которой в течение нескольких секунд или минут отсутствует расслабление мышцы. При трупном окоченении (rigor mortis), возникающем после смерти человека, значительный процент поперечных мостиков остается в связанном с актиновыми филаментами положении, что придает мышцам необычно высокую плотность.
Таким образом, функция АТФ проявляется в двух точках процесса сокращения мышцы. Во-первых, АТФ является источником энергии для сокращения. Во-вторых, АТФ отсоединяет поперечной мостик для того, чтобы повторялся цикл поперечных мостиков «замыкание-сокращение-размыкание». Активация мышечного сокращения
Процесс, при котором потенциалы действия, генерируемые на сарколемме в области нервно-мышечного синапса, активируют сокращение мышцы, называется электромеханическим сопряжением (рис. 2.29). Центральным процессом в электромеханическом сопряжении является взаимодействие ионов Са2+ с тропонином, когда изменяется положение тропомиозина, закрывающего места связывания на активном филаменте для поперечных мостиков миозина.
В состоянии покоя мышечного волокна Са2+-АТФаза саркоплазматического ретикулума активно закачивает ионы Са2+ из саркоплазмы в его цистерны, где большинство ионов находится в обратимо связанном состоянии с белком кальсеквестрином. В саркоплазме мышечного волокна концентрация ионов Са2+ снижена до уровня порядка 0,1 мкМ/л или 1(Г7 М, при котором тропонин-тропомиозиновый комплекс закрывает места связывания актина с головками поперечных мостиков миозина.
Тем не менее даже при столь низкой концентрации ионов Са2+ в саркоплазме отдельные места на концевых субдоменах актина могут осуществлять слабое электростатическое взаимодействие с миозином. В этом случае имеет место прямое связывание головки миозина с актином. Благодаря этому в состоянии покоя мышечного волокна тонкие филаменты в сарко- мере находится не в свободном, а в зафиксированном положении, что поддерживает упорядоченное пространственное расположение тонких и толстых миофиламентов между собой в саркомерах мышц.
Для того чтобы вызвать сокращение, потенциал действия мышцы, вы- зваемый двигательным нервом, распространяется по сарколемме мышечной клетки и внутрь поперечных трубочек (t-система), открывая кальциевые ионные каналы мембраны. В фазу деполяризации потенциала действия сарколеммы в области t-системы открывается L-тип потенциалзависимых кальциевых ионных каналов, которые, в свою очередь, вызывают открытие кальциевых ионных каналов саркоплазматического ретикулума. Это обусловлено тем, что L-тип потенциалзависимых кальциевых ионных каналов сарколеммы в области t-системы, или дигидропиридиновый рецептор (агонист-дигидропиридин), прямо структурно связан с рианодиновым рецептором (агонист - рианодин) или кальциевым ионным каналом саркоплазматического ретикулума. Функцией структурного взаимодействия этих
Т-трубочка мембраны мышечной клетки
Na+/K+- Na++/Ca++- Дигидропиридиновый
АТФаза обменник рецептор (Са++- канал) 

Рис. 2.29. Схема механизма электромеханического сопряжения в скелетных мышечных клетках во время начала их сокращения.
А. Механизм представляет собой последовательную цепь деполяризации сарколеммы в области Т-трубочки мышечного волокна, активации дигидропиридинового рецептора и сопряженного с ним рианодинового рецептора мембраны саркоплазматического ретикулума. Б. В результате активации обоих ионных каналов концентрация ионов кальция в цитозоле повышается за счет поступления ионов кальция из внеклеточной среды, а также из саркоплазматического ретикулума. В. Повышение концентрации ионов кальция в области сократительных белков и их связывание с субъединицей С тропонина вызывает мышечное сокращение. СПР - саркоплазматический ретикулум. Рр - рианодиновый рецептор (высвобождающий ионы кальция канал).
двух рецепторов является открытие кальциевых каналов саркоплазматического ретикулума и выход ионов Са2+ в саркоплазму. Когда концентрация ионов Са2+ в межфибриллярном пространстве, т. е. в саркоплазме между нитями актина и миозина, увеличивается до 10 мкМ/л (1(Г5 М) и более, ионы Са2+ начинают связываться с тропонином С. Активация тропонина С при связывании с ионами Са2+ вызывает смещение тропонин-тропомиози- нового комплекса примерно на 25° вокруг поверхности актинового фила- мента и погружение молекул тропомиозина вглубь между белковыми цепями актина. В результате на актиновых филаментах открываются места связывания для головок поперечных мостиков миозина и начинается цикл их сокращения. При этом начинается сокращение всей скелетной мышцы, которое будет продолжаться до тех пор, пока концентрация ионов Са2+ в саркоплазме не станет пороговой или сверхпороговой для поддержания сократительного процесса. Расслабление скелетной мышцы
Расслабление скелетной мышцы начинается только после того, как концентрация ионов Са2+ в саркоплазме мышечных клеток снижается ниже порогового уровня, т. е. ниже 10 мкМ/л, или 10-5 М. Расслабление мышцы обусловливают три процесса: 1. Отсоединение (диссоциация) ионов Са2+ от субъединицы С тропонина. 2. Диффузия ионов Са2+ в сторону саркоплазматического ретикулума. 3. Активный транспорт ионов Са2+ внутрь саркоплазматического ретикулума при участии Са2+-АТФазы.
В начальный момент расслабления длина мышцы остается постоянной. Это происходит из-за того, что первоначально происходит расслабление саркомеров, находящихся на концах мышц, в то время как центральные саркомеры остаются укороченными. Это состояние характерно лишь для начального момента мышечного расслабления, а именно фазы изометрического расслабления. Процесс расслабления во всех саркомерах усиливается после того как сила сокращения мышцы уменьшается примерно на 20- 30 %. Это обусловлено тем, что во время расслабления мышцы уменьшается подвижность тропомиозина по поверхности белковых цепей актина примерно с той же скоростью, что и сила сокращения. В результате чем меньше сила сокращения мышцы, тем меньше открыто мест связывания на актиновых филаментах для поперечных мостиков миозина Типы мышечных сокращений
Скелетные мышцы развивают сокращение под влиянием нервных импульсов, поступающих от двигательных нейронов, расположенных в передних рогах спинного мозга. Мышечные волокна, иннервируемые одним мотонейроном спинного мозга, образуют двигательную единицу. В зависимости от количества потенциалов действия, которые поступают к двигательным единицам, в мышце могут возникать несколько типов сокращения: одиночное мышечное сокращение, тетаническое, изометрическое, изотоническое и смешанное. Сокращение мышцы в целом является результатом суммации одиночных мышечных сокращений.
Одиночное мышечное сокращение. Сократительный ответ мышечного волокна или отдельной мышцы на одно раздражение называется одиночным сокращением (рис. 2.30). Одиночное сокращение имеет латентный период, фазу развития напряжения, или укорочения, и фазу удлинения, или расслабления. Длительность каждой фазы зависит от морфофункциональных свойств мышечных волокон: у быстро сокращающихся волокон глазных мышц фаза напряжения составляет 7-10 мс, а у волокон камбаловидной мышцы - 50-100 мс. Фаза укорочения в одиночном мышечном сокращении всегда меньше по времени, чем фаза расслабления. Одиночное мышечное сокращение возникает в том случае, если интервал времени между последовательными разрядами мотонейронов, вызывающими подобный тип сокращения, равен или превышает время одиночного сокращения. Так, волокна глазодвигательных мышц имеют режим одиночного сокращения при частоте импульсации мотонейрона менее 50 Гц, а волокна камбаловидной мышцы человека - при частоте менее 10 Гц. В режиме одиночного сокращения мышца способна работать длительное время без развития утомления.
Тетаническое мышечное сокращение. Тетаническим называется сокращение мышцы, при котором происходит суммация одиночных мышечных сокращений в результате ее раздражения частыми (тетаническими) стимулами. Различают зубчатый и гладкий тетанусы. Зубчатый тетанус возникает
![]()
t
t ft t t t
L
Раздражение электрическим током
Рис. 2.30. Одиночное и суммированные типы мышечного сокращения.
Одиночное сокращение мышцы (А) подразделяется на латентный период, фазу укорочения и расслабления. Суммированными типами мышечного сокращения (Б) являются зубчатый и гладкий тетанусы, которые возникают в результате суперпозиции (наложения) одиночных мышечных сокращений. Стрелками обозначены моменты нанесения электрического раздражения.
в том случае, если мышца раздражается серией электрических стимулов с частотой, при которой каждый из последующих импульсов приходится на фазу расслабления. При тетаническом сокращении мышцы фаза напряжения сменяется фазой неполного расслабления в течение всего периода раздражения. Сила сокращения, развиваемая мышцей при зубчатом тетанусе, больше, чем при одиночном мышечном сокращении. Гладкий тетанус возникает в том случае, если нерв или мышцу раздражать серией электрических импульсов с частотой, при которой каждый последующий импульс приходится на фазу напряжения. При гладком тетаническом сокращении мышца не расслабляется и в ней поддерживается постоянное напряжение. Это происходит в результате суперпозиции или наложения фазы укорочения мышцы, возникающей в мышце после каждого последующего раздражения. В результате при гладком тетанусе мышца развивает силу в 2-4 раза больше, чем при одиночном мышечном сокращении.
В разных мышцах сокращение по типу гладкого тетануса возникает при различной частоте раздражения. Гладкий тетанус быстрой глазодвигательной мышцы проявляется при частотах свыше 150-200 Гц, а у медленной камбаловидной мышцы - при частоте около 30 Гц.
Изометрическое сокращение. Если мышца развивает напряжение в условиях чрезмерной нагрузки, то ее длина не изменяется и такое сокращение называется изометрическим. Согласно теории скольжения, при изометрическом мышечном сокращении укорачивается длина саркомеров. Однако общая длина мышцы не изменяется, поскольку на такую же величину удлиняется последовательно связанный с сократительными филаментами эластический компонент мышцы.
Изотоническое сокращение. Если мышца до начала сокращения отягощена грузом, а масса груза и развиваемое мышцей напряжение во время сокращения не изменяются, то такое сокращение называется изотоническим. Во время перемещения тела в пространстве имеет место смешанный, или 4-6095
ауксотонический, или анизотонический тип сокращения, при котором мышца развивает напряжение и укорачивается. Изотонический и анизотонический типы сокращения лежат в основе динамической работы локомоторного аппарата человека.
Если внешняя нагрузка на мышцу меньше, чем развиваемое во время сокращения напряжение, то мышца укорачивается и совершается движение (концентрический тип сокращения). Если при чрезмерной внешней нагрузке мышца, напрягаясь, все же растягивается (удлиняется), то такой тип сокращения называется эксцентрическим. Типы скелетных мышечных волокон
Сокращение в одних скелетных мышцах происходит быстро, и такие мышцы быстро утомляются, а другие мышцы сокращаются медленно и так же медленно утомляются. Эта физиологическая особенность сократительной способности мышц зависит от двух типов мышечных волокон, образующих мышцу: медленно и быстро сокращающихся (рис. 2.31).
Медленные неутомляемые мышечные волокна скелетных мышц, или тип I, развивают небольшую силу при сокращении, поскольку в них мало мио- фибрилл. Низкая скорость сокращения этого типа мышечных волокон обусловлена невысокой активностью миозиновой АТФазы, медленным выходом ионов кальция из саркоплазматического ретикулума и, наконец, их медленным связыванием с тропонином. Мышечные волокна медленных двигательных единиц способны выполнять работу умеренной мощности длительное время без утомления, если хорошо развита капиллярная сеть и достаточно количество поступающего к ним 02. Например, мышцы туловища, обеспечивающие поддержание положения тела в пространстве, содержат в основном медленные мышечные волокна. Поэтому данные мышцы могут находиться в состоянии сокращения длительное время, поддерживая позу и положение тела человека в пространстве.
Быстрые мышечные волокна скелетных мышц (тип II) приспособлены для выполнения мощной, но кратковременной работы, так как богаты миофибриллами и имеют высокую скорость сокращения благодаря активности миозиновой АТФазы. Мышечные волокна этого типа мышц содержат много гликолитических ферментов (быстро освобождается большое количество энергии), в них мало митохондрий и миоглобина, они окружены небольшим количеством капилляров. Например, мышцы рук и ног имеют в своем составе преимущественно быстрые мышечные волокна, что обусловливает силу, быстроту и точность движения верхних и нижних конечностей человека.
У человека быстрые мышечные волокна подразделяют на тип НА (легко утомляемые) и ИХ (устойчивые к утомлению). Тип ИХ приспособлен для выполнения мощной работы, обладает большой аэробной выносливостью благодаря использованию энергии как аэробных, так и анаэробных процессов.
Соотношение числа медленных и быстрых двигательных единиц в одной мышце определено генетически. Например, у человека в четырехглавой мышце бедра процент медленных волокон варьирует от 40 до 98. Причем если в мышце больше медленных волокон, то такая мышца приспособлена к работе на выносливость. Если в составе мышцы выше процент быстрых волокон, то такая мышца развивает большую скорость и силу сокращения. При нарастающей работе мышц активируются первоначально медленные мышечные волокна, а затем быстрые. Это обусловлено тем, что
Alt="" />
![]()


1
1 - медленные, слабые, неутомляемые мышечные волокна. Низкий порог активации мотонейрона; 2 - промежуточный тип ДЕ; 3 - быстрые, сильные, быстроутомляемые мышечные волокна. Высокий порог активации мотонейрона.
медленные неутомляемые мышечные волокна иннервированы мотонейронами спинного мозга, которые имеют низкие пороги возбуждения и невысокую, но постоянную на протяжении длительного времени частоту генерации потенциалов действия (в среднем 6-10 Гц). Повышение силы сокращения медленных неутомляемых мышечных волокон лишь незначительно увеличивает частоту генерации потенциалов действия в мотонейронах этого типа мышечных волокон. Кроме того, нервные терминали аксонов этих мотонейронов оканчиваются на небольшой группе мышечных волокон. При силе мышечного сокращения более 20-25 % от максимальной произвольной силы активируются быстрые мышечные волокна. Это обусловлено тем, что иннервирующие их мотонейроны спинного мозга имеют высокий порог возбуждения, их частота генерации потенциалов действия нарастает по мере увеличения силы сокращения мышечных волокон, но сохраняется на постоянном уровне не продолжительное время.
Спортивная физическая нагрузка у человека не может изменить один тип мышечных волокон на другой, а также число медленных и быстрых мышечных волокон в мышце. В результате физических упражнений изменяются масса мышечной ткани, а также сила, выносливость и скоростные качества мышц тела человека.
Физиологические показатели сокращения скелетной мышцы
Основными физиологическими параметрами сокращения скелетных мышц человека являются сила, скорость развития силы и укорочения мышцы, мощность и работа
Скорость укорочения скелетной мышцы находится в обратной зависимости от величины нагрузки. Эта взаимосвязь описывается классическим уравнением Хилла. Укорочение мышечных волокон обусловливает силу мышцы во время сокращения. При этом чем больше укорочение мышцы, тем меньшую она развивает силу. Уменьшение силы мышцы по мере уменьшения ее длины во время сокращения связано с ростом концентрации ионов Са2+ в саркоплазме. Чем ближе к максимальной становится активация ионами Са2+ взаимодействия актина и миозина, тем меньшую силу развивает мышца при сокращении. Основными причинами снижения силы мышцы при ее укорочении является увеличение скорости отсоединения поперечных мостиков от актина. Этот фактор, в свою очередь, снижает сродство ионов Са2+ с местами связывания на тропонине С и высвобождает ионы Са2+ из тропонина. В результате снижается активность тропо- нин-тропомиозинового комплекса и тропомиозин блокирует места связывания на активновых филаментах для поперечных мостиков миозина. В результате снижается сила мышцы при сокращении. Скелетная мышца тела человека способна поднять груз определенной величины. Максимальная сила, которую мышца развивает при мышечном сокращении в случае, если она не в состоянии поднять груз, называется абсолютной силой мышцы. Измеряется абсолютная сила мышцы величиной минимального груза, который она не способна поднять.
Работа мышц равна произведению силы, развиваемой мышцей, на путь перемещения груза, либо механическая работа мышц равна произведению поднятого груза на величину укорочения мышцы. Между величиной нагрузки мышцы и ее укорочением нет пропорциональной зависимости. При постепенном увеличении нагрузки мышцы, которая при сокращении развивает максимальную силу, величина работы мышцы сначала растет, а затем уменьшается. В основе изменения мышечной работы при увеличении нагрузки лежат следующие физиологические процессы (рис. 2.32). При максимальном исходном растяжении мышцы (максимальная длина сарко- меров) в ней осутствует оптимальное число поперечных мостиков, которые могут сокращаться (положение 1) и развивать мышечное напряжение. При постепенном уменьшении исходного растяжения мышцы поперечные мостики миозина во все большей степени перекрываются с тонкими филамен- тами актина, что ведет к увеличению укорочения мышцы и росту работы мышцы (положение 2). Оптимальная скорость укорочения скелетной мышцы зависит от оптимальной исходной длины саркомеров перед началом сокращения, которая в среднем составляет 1,7-2,7 мкм. Однако при минимальной исходной длине саркомеров, когда миозиновые филаменты занимают всю их длину (положение 3), вновь отсутствует оптимальное число поперечных мостиков, которые могут сокращаться, развивать мышечное напряжение, а следовательно, совершать работу.
Величина мышечной работы зависит от количества мышечных волокон в мышце. При этом чем больше поперечное сечение мышцы с параллельным расположением мышечных волокон, тем большую она совершает работу. В мышцах человека не всегда мышечные волокна расположены по оси мышцы. В мышцах перистого строения, что имеет место в большинстве случаев расположения мышечных волокон относительно продольной 
Рис. 2.32. Изменение изометрического тетанического напряжения скелетной мышцы как функция от длины саркомера.
Максимальное напряжение развивается при длине мышцы L0 = 100 %. Затененная полоса показывает предел изменения длины (от 70 до 130 %), который возможен в организме, если мышцы связаны со скелетом.
оси мышцы, длина мышечных волокон нередко составляет лишь "/3- "/4 часть всей мышцы. В таких мышцах величина укорочения относительно мала, но они развивают значительную абсолютную силу и выполняют большую работу, чем мышцы с параллельным расположением мышечных волокон. Это обусловлено тем, что в перистых мышцах величина работы пропорциональна площади сечения, проходящего через все мышечные волокна, а не через анатомический поперечник мышцы в случае параллельного расположения мышечных волокон.
Наконец, мощность мышечной работы как физиологический параметр сокращения мышцы измеряется произведением величины силы мышцы на скорость укорочения мышцы при сокращении.
(одно занятие)
1. Назовите основные структурные элементы мышечного волокна, обеспечивающие его возбуждение и сокращение.
Клеточная мембрана (сарколемма), саркоплазматический ретикулум, миофибриллы.
2. Каково функциональное значение мембраны мышечного волокна в выполнении его сократительной функции?
Сарколемма является оболочкой для структурных элементов мышечного волокна, обеспечивает формирование потенциала покоя, возникновение потенциала действия и проведение возбуждения.
3. Что представляет собой миофибрилла, каково ее значение в механизме мышечного сокращения?
Миофибрилла – структура мышечного волокна, состоящая из большого числа протофибрилл (совокупность нитей актина и миозина);является сократительным элементом мышечного волокна.
4. Что представляет собой саркоплазматический ретикулум, каково его значение в механизме мышечного сокращения?
Это замкнутая система внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. Является резервуаром для хранения, выброса и обратного захвата кальция при сокращении и расслаблении мышцы.
5. Назовите структурную и функциональную единицы изолированной мышцы и двигательного аппарата в организме. Что называют двигательной единицей?
Мышечное волокно и двигательная единица соответственно. Мотонейрон с группой иннервируемых им мышечных волокон.
6. На какие группы по скорости сокращения делятся двигательные единицы, какова продолжительность их сокращения?
На быстрые и медленные. 0,01 – 0,03 с и 0,1с, соответственно.
7. Назовите группы мышц, состоящие преимущественно из быстрых или медленных мышечных волокон.
Из быстрых – некоторые мышцы глаза, мышцы пальцев рук; из медленных – дыхательные мышцы, разгибатели конечностей и спины, обеспечивающие поддержание позы.
8. Назовите функциональные отличия быстрых и медленных двигательных единиц.
У быстрых двигательных единиц скорость и сила сокращения больше, но быстрее наступает утомление, у медленных – обратные взаимоотношения.
9. Перечислите свойства мышечной ткани.
Возбудимость, проводимость, сократимость, растяжимость, эластичность.
10. Перечислите основные функции скелетных мышц.
Обеспечивают все виды двигательной активности, поддержание определенной позы, дыхательную функцию, жевание, выработку тепла, способствуют движению крови и лимфы по сосудам к сердцу.
11. Что называют сократимостью мышцы? Что является непосредственной причиной сокращения (укорочения) мышцы?
Способность мышечной ткани изменять длину или напряжение. Скольжение нитей актина вдоль нитей миозина навстречу друг другу.
12. Почему потенциал действия считается инициатором мышечного сокращения? Дайте соответствующие пояснения.
Потенциал действия повышает проницаемость саркоплазматического ретикулума, что обеспечивает выход из него ионов кальция, необходимых для запуска процесса сокращения мышцы.
13. Нарисуйте потенциал действия скелетной мышцы, полученный при внутриклеточном отведении. Укажите его амплитуду в мВ.
14. Нарисуйте, сопоставив во времени, потенциал действия и цикл одиночного сокращения скелетной мышцы. Назовите фазы сокращения мышцы.
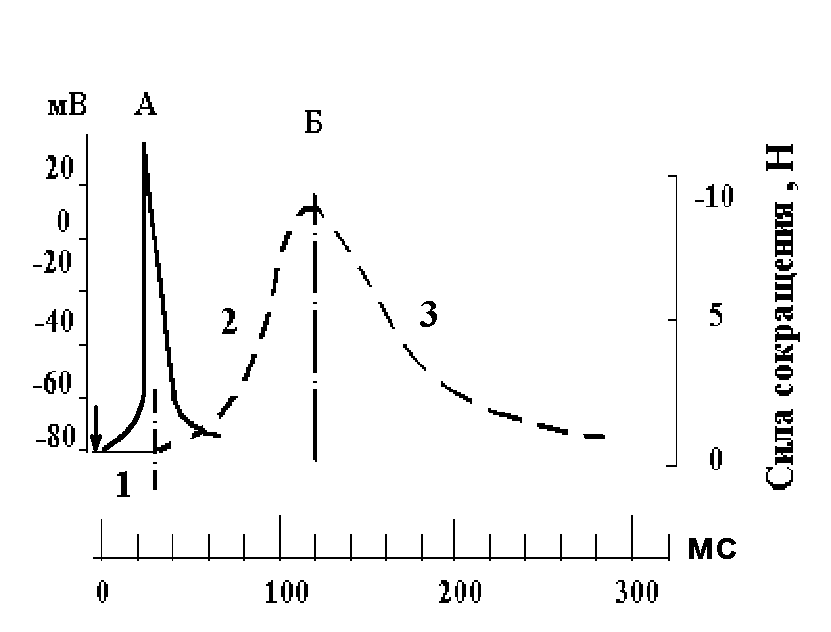 А
– потенциал действия; Б – изометрическое
сокращение мышцы: 1 – латентный период;
2 – фаза напряжения; 3 – фаза расслабления;
– момент нанесения раздражения.
А
– потенциал действия; Б – изометрическое
сокращение мышцы: 1 – латентный период;
2 – фаза напряжения; 3 – фаза расслабления;
– момент нанесения раздражения.
15. Опишите кратко роль ионов кальция в механизме мышечного сокращения.
Ионы кальция взаимодействуют с белковым комплексом тропонин-тропомиозин, что ведет к освобождению активных участков на нитях актина и зацеплению за них головок миозиновых мостиков.
16. На какие процессы, обеспечивающие сокращение мышцы, расходуется энергия АТФ?
На взаимодействие актиновых и миозиновых нитей, обеспечивающее их скольжение относительно друг друга (укорочение) и работу ионных насосов.
17. Опишите последовательно процессы, обеспечивающие освобождение энергии АТФ при мышечном сокращении.
Контакт головок миозина с нитями актина – активация АТФ-азы миозина в присутствии ионов магния – расщепление АТФ – выделение энергии.
18. Что является непосредственной причиной скольжения нитей актина и миозина, обеспечивающего мышечное сокращение? Почему?
"Сгибание" миозиновых мостиков. Потому что в этот момент они «зацеплены» своими головками за активные участки нитей актина.
19. Активным (с затратой энергии АТФ) или пассивным (без затраты энергии АТФ) является процесс расслабления мышцы?
Расслабление мышцы обеспечивают как активные, так и пассивные процессы.
20. Какой из процессов, обеспечивающих мышечное расслабление, является активным, какой – пассивным?
Активным (с затратой энергии АТФ) является процесс переноса ионов кальция в саркоплазматический ретикулум, пассивным – скольжение нитей актина и миозина, ведущее к уменьшению зон их взаимного перекрытия.
21. Что является причиной скольжения нитей актина и миозина относительно друг друга при расслаблении мышцы.
Эластические свойства самой мышцы и сухожилия, растянутых при сокращении мышцы и масса органа (сила тяжести).
22. Назовите две фазы теплообразования в мышцах. Каким периодам мышечного сокращения соответствует первая из них?
Первая фаза – начальное теплообразование, соответствует возбуждению, укорочению и расслаблению мышцы, вторая – восстановительное (запаздывающее) теплообразование.
23. На какие процессы расходуется энергия восстановительного теплообразования в мышцах?
На ресинтез АТФ и работу ионных помп, обеспечивающих перенос ионов Na + , K + и Ca 2+ .
24. Назовите источники энергии, обеспечивающие ресинтез АТФ.
Расщепление креатинфосфата, анаэробный гликолиз; аэробное окисление углеводов и жирных кислот, т. е. через окислительное фосфорилирование.
25. На что расходуется энергия (укажите распределение в процентах), освобождаемая при мышечном сокращении? Каков КПД мышцы?
50% энергии расходуется на сокращение и расслабление (из них: 25% – на механическую деятельность [это КПД мышцы], 25% – на работу ионных помп); 50% тепла выделяется в окружающую среду.
26. Назовите типы сокращения скелетных мышц в зависимости от условий сокращения и от характера раздражения.
В зависимости от условий сокращения различают изометрическое и изотоническое сокращения. В зависимости от характера раздражения различают одиночное и тетаническое сокращения.
27. Назовите три фазы одиночного мышечного сокращения. Какой основной процесс происходит в первую фазу?
Латентный период, период укорочения и расслабления. Возбуждение.
28. Какие факторы влияют на силу одиночного мышечного сокращения?
Степень предварительного растяжения мышцы и сила ее раздражения.
29. Почему увеличение силы раздражения мышцы увеличивает силу ее сокращения?
Вследствие увеличения числа сокращающихся волокон. В возбуждение дополнительно вовлекаются волокна, которые при слабом раздражении не возбуждались из-за более низкой их возбудимости или более глубокого расположения в мышце.
30. Почему предварительное умеренное растяжение изолированной мышцы увеличивает силу ее сокращения при одиночном раздражении?
В результате того, что в умеренно растянутой мышце увеличивается как пассивное напряжение упругих элементов, так и активная сила сокращения вследствие увеличения зон взаимодействия нитей актина и миозина.
31. До какой степени необходимо растянуть изолированную мышцу, чтобы сила ее активного сокращения в изометрическом режиме была максимальной при одиночном раздражении? Почему?
До ее длины в состоянии покоя in situ, что обеспечивает максимальную зону контакта между нитями актина и миозина, и следовательно, максимальное число точек взаимодействия миозиновых мостиков с нитями актина.
32. Как и почему будет изменяться сила активного сокращения мышцы, если ее предварительно растянуть более ее длины в состоянии покоя in situ, а затем, перед каждым последующим раздражением увеличивать степень ее растяжения?
Будет уменьшаться вплоть до нуля в результате уменьшения зон взаимного перекрытия нитей актина и миозина (уменьшается число точек зацепления миозиновых мостиков с нитями актина, вплоть до полного их отсутствия).
33. Что называют тетаническим сокращением мышцы? Какое явление лежит в основе механизма тетануса?
Слитное, длительное сокращение скелетной мышцы, возникающее в ответ на ритмическое раздражение. Явление суммации мышечных сокращений.
34. Что называют суммацией мышечных сокращений?
Увеличение силы (или амплитуды) и длительности сокращения мышцы под действием ее повторного раздражения в период предыдущего сокращения.
35. За счет чего увеличивается амплитуда мышечного сокращения при суммации в изотоническом режиме? Объясните механизм.
За счет дополнительного скольжения нитей актина и миозина относительно друг друга в результате увеличения зон зацепления миозиновых мостиков под влиянием дополнительного выхода ионов Са 2+ из саркоплазматического ретикулума.
36. Почему при суммации мышечных сокращений в изометрическом режиме возможно дополнительное скольжения нитей актина относительно миозина?
Потому что мышца обладает растяжимостью, что делает возможным дополнительное укорочение саркомеров.
37. При каких условиях раздражения скелетной мышцы вместо одиночных сокращений возникает тетанус? Какие виды тетануса Вам известны?
При ритмическом раздражении с интервалами между импульсами меньшими, чем период одиночного мышечного сокращения. Зубчатый и гладкий.
38. В какую фазу одиночного сокращения должно попасть каждое последующее раздражение, чтобы возник зубчатый или гладкий тетанус? Какие факторы влияют на высоту гладкого тетануса изолированной мышцы?
В фазу расслабления мышцы или в фазу укорочения (напряжения) мышцы соответственно. Степень предварительного растяжения мышцы, сила и частота ее раздражения.
39. Какова зависимость высоты гладкого тетануса от частоты раздражения мышцы (в динамике)?
С увеличением частоты раздражения до известного предела величина тетанического сокращения нарастает, а затем уменьшается, вплоть до полного расслабления мышцы.
40. Какую частоту раздражения мышцы называют оптимальной, какую – пессимальной?
Оптимальная частота – при которой гладкий тетанус наиболее высокий и устойчивый, пессимальная – высокая частота, превышающая лабильность мышц, при которой она расслабляется.
41. Почему при оптимальной частоте раздражения изолированной мышцы тетанус наиболее высокий и устойчивый, а при пессимальной частоте раздражения мышца расслабляется?
Потому что при оптимальной частоте раздражения каждый последующий стимул попадает в фазу экзальтации, а при пессимальной – в фазу абсолютной рефрактерности.
42. В каком звене нервно-мышечного препарата (нерв-синапс-мышца) и почему при ритмическом раздражении нерва развивается пессимальное торможение Н. Е. Введенского?
В мионевральном синапсе, так как его лабильность наименьшая.
43. С помощью какого приема, можно доказать, что пессимум не связан с утомлением мионеврального синапса?
Уменьшение частоты раздражения до оптимальной сразу же ведет к восстановлению исходного гладкого тетануса.
44. Синхронно или асинхронно сокращаются отдельные мышечные волокна в естественных условиях? За счет каких механических эффектов увеличивается сила сокращения скелетной мышцы в естественных условиях?
Асинхронно. За счет вовлечения в реакцию большего числа двигательных единиц, увеличения степени синхронизации их возбуждения, дополнительного скольжения нитей актина и миозина относительно друг друга в каждой миофибрилле.
45. Подчиняется ли двигательная единица закону "все или ничего"? Почему?
Подчиняется, так как импульсы по ветвлениям аксона подходят одновременно ко всем мышечным волокнам двигательной единицы, и они сокращаются синхронно, т. е. двигательная единица функционирует как единое целое.
46. В каких условиях отдельные двигательные единицы одного нейронного пула возбуждаются синхронно, в каких – асинхронно?
При умеренных мышечных нагрузках – асинхронно, т. е. независимо друг от друга, при чрезмерных усилиях – синхронно.
47. Почему асинхронное возбуждение отдельных двигательных единиц в естественных условиях дает плавное тетаническое сокращение скелетных мышц?
Вследствие суммации сокращений большого числа асинхронно сокращающихся мышечных волокон различных двигательных единиц.
48. В каких отделах центральной нервной системы находятся мотонейроны, аксоны которых иннервируют скелетные мышцы?
В передних рогах спинного мозга, в продолговатом мозгу, в мосту и среднем мозге.
49. Что называют тонусом скелетных мышц, развивается ли при этом их утомление, велик ли расход энергии?
Постоянное слабое напряжение (сокращение) скелетных мышц, поддерживаемое редкими импульсами из центральной нервной системы и осуществляемое с малым расходом энергии без признаков утомления.
50. Какова зависимость работы изолированной скелетной мышцы от величины нагрузки?
С увеличением нагрузки работа мышцы сначала возрастает, а затем уменьшается вплоть до нуля при чрезмерно сильных нагрузках, когда мышца не в состоянии поднять груз.
51. Сформулируйте правило "средних нагрузок". Как и почему изменится работоспособность скелетной мышцы при увеличении частоты ее сокращений?
Работа мышцы максимальна при средних нагрузках. С увеличением частоты сокращений работоспособность сначала возрастает, а при превышении оптимальной частоты работоспособность уменьшается, т. к. быстрее развивается утомление.
52. Что называют утомлением мышцы? Чем оно объясняется?
Временное понижение работоспособности мышцы, наступающее в результате работы и исчезающее после отдыха. Накоплением продуктов обмена веществ, постепенным истощением энергетических запасов.
53. В условиях целого организма или в изолированной мышце утомление наступает медленнее? Почему?
В условиях целого организма, т. к. мышца постоянно снабжается кровью: получает питательные вещества и кислород и освобождается от продуктов метаболизма.
54. Где в условиях целого организма утомление наступает раньше: в центральной нервной системе, в нервно-мышечном синапсе или в самой мышце? Что такое активный отдых?
В центральной нервной системе. Восстановление работоспособности утомленных мышц при двигательной активности других мышц.
55. Как доказать в эксперименте, что в изолированной мышце утомление связано с накоплением продуктов обмена?
Изолированную мышцу поместить в раствор Рингера и длительным раздражением вызвать ее утомление, после замены раствора – работоспособность мышцы на некоторое время восстанавливается.
56. Перечислите структурные особенности гладкой мышцы.
Нерегулярное расположение нитей актина и миозина, вследствие чего отсутствует поперечная исчерченность, слабое развитие саркоплазматического ретикулума, наличие нексусов между мышечными волокнами.
57. Перечислите особенности потенциала покоя и потенциала действия гладкой мышцы по сравнению с таковыми поперечнополосатой мышцы.
Величина потенциала покоя гладкой мышцы меньше (30 – 50 мВ), наблюдается спонтанная деполяризация. Потенциалы действия бывают пикообразными и платообразными, более продолжительны – до 0, 5 с, деполяризация обеспечивается, главным образом, с помощью кальция и частично посредством натрия.
58. Назовите функциональные особенности гладкой мышцы по сравнению со скелетной.
Гладкой мышце присущи: автоматия, пластичность, более продолжительное сокращение (от нескольких секунд до 1 мин).
59. Что такое пластичность гладких мышц, каково ее значение для функционирования внутренних полых органов?
Способность длительно сохранять приданную растяжением длину без изменения напряжения. Благодаря пластичности гладкой мышцы давление в полых органах почти не меняется при медленном их наполнения.
60. Что является функциональной единицей гладкой мышцы? Почему?
Пучок мышечных волокон, заключенный в соединительнотканную оболочку, в пределах которого возбуждение передается от одного волокна к другому, охватывая весь пучок одновременно.
1. Какими свойствами обладает скелетная мышца плода к моменту рождения? Как изменяется упругость, прочность и эластичность мышц с возрастом?
Возбудимостью, проводимостью, сократимостью, эластичностью и растяжимостью, т. е. всеми свойствами мышцы взрослого. Упругость и прочность увеличиваются, эластичность уменьшается.
2. Как изменяются в онтогенезе возбудимость, проводимость, сократимость, утомляемость, быстрота сокращения и расслабления, лабильность скелетной мышцы?
Показатели всех перечисленных свойств увеличиваются.
3. Каковы соотношения силы мышц мальчиков и девочек в период от 7 до 8 лет, в возрасте 10 – 12 лет и 15 – 18 лет?
До 7 – 8 летнего возраста сила их мышц одинакова, в 10 – 12 лет – больше у девочек, в 15 – 18 лет – больше у мальчиков.
4. Укажите величину мембранного потенциала мышечного волокна новорожденного ребенка и взрослого человека. С чем связано это различие?
У новорожденных 20 – 40 мВ, у взрослых 80 –90 мВ. У новорожденных больше ионная проницаемость мембраны мышечного волокна, поэтому больше утечка ионов, меньше градиент их концентрации.
5. Перечислите отличия потенциала действия мышечного волокна новорожденного от такового у взрослого.
Меньшая амплитуда, большая продолжительность, часто отсутствует инверсия.
6. Увеличивается или уменьшается скорость проведения возбуждения по мышечному волокну с возрастом? Перечислите факторы, обеспечивающие это изменение.
Увеличивается. Увеличение мембранного потенциала, потенциала действия, а также диаметра мышечного волокна.
7. Почему увеличение амплитуды потенциала действия мышечного волокна в процессе роста организма увеличивает скорость проведения возбуждения?
Больший потенциал действия быстрее вызывает возбуждение соседнего участка мышечного волокна.
8. Почему увеличение диаметра мышечного волокна в процессе роста организма увеличивает скорость проведения возбуждения?
Увеличение толщины мышечного волокна ведет к уменьшению продольного сопротивления ионному току в миоплазме.
9. Перечислите особенности сокращения мышц новорожденного. Разделяются ли мышцы новорожденного на быстрые и медленные?
Относительная длительность одиночного сокращения (фазы укорочения и расслабления), тонические сокращения, без признаков пессимального торможения при большой частоте раздражения. Не подразделяются.
10. Как изменяется эффективность отдыха (становится больше или меньше) после физического утомления у детей разного возраста: 7 – 12 лет, 13 – 15 лет и в 16 – 18 лет?
В 7 – 12 лет эффективность отдыха наибольшая, в 13 – 15 лет резко падает, в 16 – 18 лет несколько увеличивается.
11. В каком возрасте наблюдается максимальная выносливость к физическим нагрузкам?
В возрасте 20 – 29 лет.
12. Что представляет собой незрелый (примитивный) нервно-мышечный синапс новорожденного? В чем заключается его функциональная особенность, к какому возрасту заканчивается его созревание?
На поверхности миотрубки выемка, в которой расположено окончание аксона. Значительно большая синаптическая задержка (в 7 – 10 раз больше, чем у взрослых). К 7 – 8 годам.
13. В чем выражается созревание терминальных ветвлений аксона мотонейрона? Какое значение имеет этот процесс?
В увеличении терминальных ветвлений аксона и содержания в них ацетилхолина. Это ведет к увеличению выхода медиатора в синаптическую щель при поступлении импульса к нервному окончанию и увеличению амплитуд ПКП.
14. В чем выражается созревание постсинаптической мембраны?
В увеличении плотности холинорецепторов на ней, образовании складок, увеличении потенциала концевой пластинки, появлении в ней холинэстеразы.
15. Как и почему изменяется синаптическая задержка в нервно-мышечном синапсе в процессе созревания?
Уменьшается вследствие увеличения скорости освобождения ацетилхолина из пресинаптического окончания на каждый нервный импульс, увеличения потенциала концевой пластинки и появления в ней холинэстеразы.
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯХАРАКТЕРИСТИКА СКЕЛЕТНОЙ
МЫШЦЫ И МЕХАНИЗМ ЕЕ
СОКРАЩЕНИЯСтруктурной единицей скелетной мышцы
является мышечное волокно - сильно вытянутая
многоядерная клетка.
Длина мышечного волокна зависит от размеров
мышцы и составляет от нескольких миллиметров
до нескольких сантиметров. Толщина волокна
варьирует от (10-100 мкм).
Типы мышц
В организме человека существует три типа
мышц:
скелетные, сердечные (миокард) и гладкие.
При микроскопическом исследовании в
скелетных и сердечной мышцах
обнаруживается исчерченностъ, поэтому их
называют поперечнополосатыми мышцами.Скелетные мышцы прикреплены в основном к
костям, что и обусловило их название.
Сокращение скелетных мышц инициируется
нервными
импульсами
и
подчиняется
сознательному
контролю,
т.е.
осуществляется произвольно.
Сокращение гладких мышц инициируется
импульсами, некоторыми гормонами и не
зависит от воли человека.Мышечное волокно окружено двухслойной
липопротеидной электровозбудимой мембраной сарколеммой,
которая
покрыта
сетью
коллагеновых волокон, придающих ей прочность и
эластичность.
В скелетных мышцах различают несколько типов
мышечных волокон: медленносокращающиеся
(МС) или красные и быстросокращающиеся
(БС) или белые.
Молекулярный механизм сокращения.
Скелетные мышцы содержат сократительные
белки:
актин
и
миозин.
Механизм
их
взаимодействия во время элементарного акта
мышечного
сокращения
объясняет
теория
скользящих нитей, разработанная Хасли и
Хансоном.
Строение мышечного волокна
Сарколемма – плазматическая мембрана покрывающаямышечное волокно (соединяется с сухожилием, которое
прикрепляет мышцу к кости; сухожилие передает усилие
производимое мышечными волокнами кости и таким
образом
осуществляется
движение).
Сарколемма
обладает избирательной проницаемостью для различных
веществ и имеет транспортные системы, с помощью
которых поддерживается разная концентрация ионов
Na+, К+, а также Сl- внутри клетки и в межклеточной
жидкости, что приводит к возникновению на ее
поверхности мембранного потенциала - необходимого
условия возникновения возбуждения мышечного волокна.
Саркоплазма
–
желатиноподобная
жидкость,
заполняющая
промежутки
между
миофибриллами
(содержит
растворенные
белки,
микроэлементы,
гликоген, миоглобин, жиры, органеллы). Около 80%
объема волокна занимают длинные сократительные нити
- миофибриллы.Система поперечных трубочек. Это сеть Т –
трубочек (поперечные), является продолжением
сарколеммы; они взаимосоединяются проходя
среди миофибрилл. Обеспечивают быструю
передачу нервных импульсов (распространение
возбуждения) внутрь клетки к отдельным
миофибриллам.
Саркоплазматический ретикулум (СР) – сеть
продольных трубочек, расположены параллельно
миофибриллам; это место депонирования Са2+,
который необходим для обеспечения процесса
мышечного сокращения.
Сократительные белки актин и миозин образуют
в миофибриллах тонкие и
толстые
миофиламенты.
Они
располагаются
параллельно друг другу внутри мышечной клетки
Миофибриллы
представляют
собой
сократимые элементы мышечного волокна пучки «нитей» (филаментов).Структура миофибриллы:
1. Перегородки – называемые Z - пластинками,
разделяют их на саркомеры.
Структура саркомера:
В них видна последовательность регулярно
чередующихся поперечных светлых и темных
полос,
которая
обусловлена
особым
взаиморасположением
актиновых
и
миозиновых
филаментов
(поперечная
полосатость).
Середину саркомера занимают «толстые» нити
миозина. (А – диск темный)
На
обоих концах саркомера находятся
«тонкие» нити актина. (I- диск светлый)Актиновые нити прикрепляются к Z –
пластинкам, сами Z – пластинки
ограничивают саркомер.
В покоящейся мышце концы тонких и
толстых
филаментов
лишь
слабо
перекрываются на границе между А и Iдисками.
Н – зона (светлее) в которой нет
перекрывания
нитей
(здесь
располагаются только миозиновые нити),
находится в диске А.
М - линия находится в центре саркомера
– место удержания толстых нитей
(построена из опорных белков.)
Теория скользящих нитей.
Укорочение саркомера:Мышца сокращается в результате укорочения множества
последовательно соединенных саркомеров в
миофибриллах.
Во время сокращения тонкие актиновые филаменты
скользят вдоль толстых миозиновых, двигаясь между ними
к середине их пучка и саркомера.
Основное положение теории скользящих нитей:
Во время сокращения мышцы, сами актиновые и
миозиновые нити не укорачиваются (ширина А – диска
всегда остается постоянной, тогда как I- диски и Н – зоны
при сокращении сужаются).
Длинна нитей не меняется при растяжении мышцы (тонкие
филаменты вытягиваются из промежутков между толстыми
нитями, так что степень перекрывания их пучков
уменьшается).
10. Работа поперечных мостиков.
Движение головок создает объединенное усилие,как бы «гребок», продвигающий актиновые нити к
середине саркомера. Только за счет ритмичных
отделений и повторных прикреплений миозиновых
головок актиновая нить может подтягиваться к
середине саркомера.
При расслаблении мышцы миозиновые головки
отделяются от актиновых нитей.
Так как актиновые и миозиновые нити могут легко
скользить друг относительно друга, сопротивление
расслабленных мышц растяжению очень низкое.
Удлинение мышцы во время расслабления носит
пассивный характер.
11. Преобразование химической энергии в механическую.
АТФ – непосредственный источник энергии длясокращения.
При сокращении мышцы АТФ расщепляется на
АДФ и фосфат.
Ритмическая активность поперечных мостиков, т.
е. циклы их прикрепления к актину и отсоединения
от него, обеспечивающие мышечное сокращение,
возможны только при гидролизе АТФ, а
соответственно и при активации АТФазы, которая
непосредственно участвует в расщеплении АТФ на
АДФ и фосфат.
12. Молекулярный механизм мышечного сокращения.
Сокращение запускается нервным импульсом. При этом всинапсе - месте контакта нервного окончания с
сарколеммой выделяется медиатор (нейропередатчик) ацетилхолин.
Ацетилхолин (Ах) вызывает изменение проницаемости
мембраны для некоторых ионов, что в свою очередь
приводит к возникновению ионных токов и сопровождается
деполяризацией мембраны. В следствии чего, на ее
поверхности возникает потенциал действия или она
возбуждается.
Потенциал
действия
(возбуждение)
распространяется вглубь волокна через Т-системы.
Нервный импульс вызывает изменение проницаемости
мембраны саркоплазматического ретикулума и приводит к
освобождению
ионов
Са2+
из
пузырьков
саркоплазматического ретикулума.
13. Электромеханическое сопряжение
Передача команды к сокращению отвозбужденной клеточной мембраны к
миофибриллам
в
глубине
клетки
(электромеханическое
сопряжение)
включает
в
себя
несколько
последовательных процессов, ключевую
роль в которых играют ионы Са2+.
14.
1. Электромеханическое сопряжение происходитпосредством распространения потенциала
действия по мембранам поперечной системы
внутрь клетки, потом возбуждение проходит на
продольную систему (ЭПР) и вызывает
высвобождение депонированного в мышечной
клетке Са2+ во внутриклеточное пространство,
которое окружает миофибриллы. Это и приводит к
сокращению
2. Са2+ удаляется из внутриклеточного пространства
в депо (каналы ЭПР) за счет работы кальциевых
насосов на мембранах ЭПР.
3. Только за счет электрической передачи по
поперечной системе, возможна быстрая
мобилизация запасов кальция в глубине волокна, и
только этим можно объяснить очень короткий
латентный период между стимулом и
сокращением.
15.
Функциональная роль АТФ:- в покоящейся мышце - препятствует соединению
актиновых нитей с миозиновыми;
- в процессе сокращения мышцы - поставляет
необходимую энергию для движения тонких нитей
относительно толстых, что приводит к укорочению
мышцы или развитию напряжения;
- в процессе расслабления - обеспечивает энергией
активный транспорт Са2+ в ретикулум.
16. Типы мышечных сокращений. Оптимум и пессимум мышечного сокращения
В зависимости от изменения длины мышечного волокнавыделяют два типа его сокращения - изометрическое и
изотоническое.
Мышечное сокращение при котором длина мышцы
уменьшается по мере развиваемой ею силы, называется
ауксотоническим.
Максимальная сила при ауксотонических экспериментальных
условиях (с растяжимой упругой связью между мышцей и
датчиком силы) называется максимумом ауксотонического
сокращения. Она гораздо меньше силы, которую развивает
мышца при постоянной длине, т.е. при изометрическом
сокращении.
Сокращение мышцы, при котором ее волокна укорачиваются
при неизменном напряжении, называется изотоническим.
Сокращение мышцы, при котором ее напряжение возрастает
а длина мышечных волокон остается неизменной,
называется изометрическим
17.
Мышечная работа равна произведениюрасстояния (укорочения мышцы) на вес груза,
который поднимает мышца.
При изотонической тетанической активации
мышцы от нагрузки зависит величина укорочения и
скорость укорочения мышцы.
Чем меньше нагрузка, тем больше укорочений в
единицу времени. Ненагруженная мышца
укорачивается с максимальной скоростью, которая
зависит от типа мышечных волокон.
Мощность мышцы равна произведению
развиваемой ею силы на скорость укорочения
18.
Расслабленная мышца, сохраняющая «длину покоя» за счетфиксации обоих ее концов, не развивает силу, которая
передавалась бы на датчик. Но если потянуть за один ее
конец, чтобы волокна растянулись, в ней возникает
пассивное напряжение. Таким образом, мышца в состоянии
покоя упруга. Модуль упругости покоящейся мышцы с
растяжением возрастает. Эта упругость обусловлена главным
образом растяжимыми структурами, которые располагаются
параллельно
относительно
растяжимых
миофибрилл
(«параллельная
упругость»)
.
Миофибриллы
в
расслабленном состоянии практически не оказывают
сопротивления растяжению; актиновые и миозиновые нити, не
связанные
поперечными
мостиками,
легко
скользят
относительно друг друга. Степень предварительного
растяжения определяет величину пассивного напряжения
покоящейся мышцы и величину дополнительной силы,
которую может развить мышца в случае активации при данной
длине.
19.
Пиковое усилие при таких условиях называетсямаксимумом изометрического сокращения.
При сильном растяжении мышцы, сила сокращения
уменьшается т. к. нити актина вытянуты из
миозиновых пучков и соответственно, меньше зона
перекрывания этих нитей и возможность
формирования поперечных мостиков.
При очень сильном растяжении мышцы, когда
актиновые и миозиновые нити перестают
перекрываться, миофибриллы не способны
развивать силу. Это доказывает, что мышечная сила
представляет собой результат взаимодействия
актиновых и миозиновых филаментов (т. е.
образования между ними поперечных мостиков).
В естественных условиях сокращения мышц
являются смешанными - мышца обычно не только
укорачивается, но изменяется и ее напряжение.
20.
В зависимости от длительности выделяютодиночное и тетаническое сокращения мышцы.
Одиночное сокращение мышцы в эксперименте
вызывают одиночным раздражением электрическим
током. В изотоническом режиме одиночное
сокращение начинается через короткий скрытый
(латентный) период, далее следует фаза подъема
(фаза укорочения), затем фаза спада (фаза
расслабления) (рис. 1). Обычно мышца
укорачивается на 5-10% исходной длины.
Длительность ПД мышечных волокон также
варьирует и составляет 5-10 мс с учетом замедления
фазы реполяризации.
Мышечное волокно подчиняется закону «все или
ничего», т.е. отвечает на пороговое и
сверхпороговое раздражение одинаковым по
величине одиночным сокращением.
21.
Сокращение целой мышцы зависит:1. от силы раздражителя при непосредственном раздражении
мышцы
2. от числа нервных импульсов, поступающих к мышце при
раздражении нерва.
Увеличение силы раздражителя ведет к увеличению числа
сокращающихся мышечных волокон.
Подобный эффект наблюдается и в естественных условиях - с
увеличением числа возбужденных нервных волокон и частоты
импульсов (к мышце поступает больше нервных импульсов ПД) увеличивается число сокращающихся мышечных волокон.
При одиночных сокращениях мышца утомляется
незначительно.
Тетаническое сокращение - это слитное длительное
сокращение скелетной мышцы. В его основе лежит явление
суммации одиночных мышечных сокращений.
Кривая одиночного
сокращения икроножной
мышцы лягушки:
1-латентный период,
2- фаза укорочения,
22.
При нанесении на мышечное волокно илинепосредственно
на
мышцу
двух
быстро
следующих друг за другом раздражений,
возникающее
сокращение
имеет
большую
амплитуду и длительность. При этом нити актина и
миозина дополнительно скользят друг относительно
друга. В сокращение могут вовлекаться ранее не
сокращавшиеся мышечные волокна, если первый
стимул вызвал у них подпороговую деполяризацию,
а второй увеличивает ее до критической величины.
Суммация сокращений при повторном раздражении
мышцы или поступлении к ней ПД возникает только
в том случае, когда закончен рефрактерный период
(после исчезновения ПД мышечного волокна).
23.
При поступлении импульсов к мышце во время еерасслабления возникает зубчатый тетанус, во
время укорочения - гладкий тетанус (рис.).
Амплитуда тетануса больше величины
максимального одиночного сокращения мышцы.
Напряжение, развиваемое мышечными волокнами
при гладком тетанусе, обычно в 2-4 раза больше,
чем при одиночном сокращении, однако мышца
быстрее утомляется. Мышечные волокна не
успевают восстановить энергетические ресурсы,
израсходованные во время сокращения.
Амплитуда гладкого тетануса увеличивается с
возрастанием частоты стимуляции нерва. При
некоторой (оптимальной) частоте стимуляции
амплитуда гладкого тетануса наибольшая (оптимум частоты раздражения)
24.
Рис. Сокращения икроножной мышцы лягушки приувеличении частоты раздражения седалищного нерва
(ст/с - стимулов в секунду): а - одиночное сокращение;
б-д - накладывание волн сокращения друг на друга и
образование разных видов тетанического сокращения.
При частоте 120 ст/с - пессимальный эффект
(расслабление мышцы во время стимуляции) – е
25.
При чрезмерно частой стимуляции нерва (более 100имп/с) мышца расслабляется вследствие блока
проведения возбуждения в нервно-мышечных
синапсах - пессимум Введенского (пессимум
частоты раздражения). Пессимум Введенского можно
получить и при прямом, но более частом раздражении
мышцы (более 200 имп/с) . Пессимум Введенского не
является результатом утомления мышцы или истощения медиатора в синапсе, что доказывается фактом
возобновления сокращения мышцы сразу же после
уменьшения частоты раздражения. Торможение
развивается в нервно-мышечном синапсе при
раздражении нерва.
В естественных условиях мышечные волокна
сокращаются в режиме зубчатого тетануса или
даже одиночных последовательных сокращений.
26.
Однако форма сокращения мышцы в целомнапоминает гладкий тетанус.
Причины
этого
асинхронность
разрядов
мотонейронов и асинхронность сократительной
реакции отдельных мышечных волокон, вовлечение
в сокращение большого их количества, вследствие
чего мышца плавно сокращается и плавно
расслабляется, может длительно находиться в
сокращенном состоянии за счет чередования
сокращений множества мышечных волокон. При
этом мышечные волокна каждой двигательной
единицы сокращаются синхронно.
27.
Функциональная единица мышцы –двигательная единица
Понятия. Иннервация скелетных мышечных волокон
осуществляется мотонейронами спинного мозга или
мозгового ствола. Один мотонейрон веточками своего
аксона иннервирует несколько мышечных волокон.
Совокупность мотонейрона и иннервируемых им
мышечных волокон называют двигательной
(нейромоторной) единицей. Число мышечных
волокон двигательной единицы варьирует в широких
пределах в разных мышцах. Двигательные единицы
невелики в мышцах, приспособленных для быстрых
движений, от нескольких мышечных волокон до
нескольких десятков их (мышцы пальцев, глаза,
языка). Наоборот, в мышцах, осуществляющих
медленные движения (поддержание позы мышцами
туловища), двигательные единицы велики и включают
сотни и тысячи мышечных волокон
28.
Присокращении
мышцы
в
натуральных
(естественных) условиях можно зарегистрировать
ее электрическую активность (электромиограмму ЭМГ) с помощью игольчатых или накожных электродов. В абсолютно расслабленной мышце
электрическая активность почти отсутствует. При
небольшом
напряжении,
например
при
поддержании
позы,
двигательные
единицы
разряжаются с небольшой частотой (5-10 имп/с),
при большом напряжении частота импульсации
повышается в среднем до 20-30 имп/с. ЭМГ позволяет судить о функциональной способности
нейромоторных единиц. С функциональной точки
зрения двигательные единицы разделяют на
медленные и быстрые.
29.
мотонейроны и медленные мышечные волокна (красные).Медленные мотонейроны, как правило, низкопороговые, так
как обычно это малые мотонейроны. Устойчивый уровень
импульсации у медленных мотонейронов наблюдается уже
при очень слабых статических сокращениях мышц, при
поддержании позы. Медленные мотонейроны способны
поддерживать длительный разряд без заметного снижения
частоты импульсации на протяжении длительного времени.
Поэтому их называют малоутомляемыми или
неутомляемыми мотонейронами. В окружении медленных
мышечных волокон богатая капиллярная сеть, позволяющая
получать большое количество кислорода из крови.
Повышенное содержание миоглобина облегчает транспорт
кислорода в мышечных клетках к митохондриям. Миоглобин
обусловливает красный цвет этих волокон. Кроме того,
волокна содержат большое количество митохондрий и
субстратов окисления - жиров. Все это обусловливает использование медленными мышечными волокнами более
эффективного аэробного окислительного пути
30.
Быстрые двигательные единицы состоят избыстрых мотонейронов и быстрых мышечных
волокон. Быстрые высокопороговые мотонейроны
включаются в активность только для обеспечения
относительно больших по силе статических и
динамических сокращений мышц, а также в начале
любых сокращений, чтобы увеличить скорость
нарастания напряжения мышцы или сообщить
движущейся части тела необходимое ускорение. Чем
больше скорость и сила движений, т. е. чем больше
мощность сократительного акта, тем больше участие
быстрых двигательных единиц. Быстрые
мотонейроны относятся к утомляемым - они не
способны к длительному поддержанию
высокочастотного разряда
31.
Быстрые мышечные волокна (белые мышечныеволокна) более толстые, содержат больше
миофибрилл, обладают большей силой, чем
медленные волокна. Эти волокна окружает меньше
капилляров, в клетках меньше митохондрий,
миоглобина и жиров. Активность окислительных
ферментов в быстрых волокнах ниже, чем в
медленных, однако активность гликолитических
ферментов, запасы гликогена выше. Эти волокна не
обладают большой выносливостью и более
приспособлены для мощных, но относительно
кратковременных сокращений. Активность быстрых
волокон имеет значение для выполнения
кратковременной высокоинтенсивной работы,
например бега на короткие дистанции
32.
Скорость сокращения мышечных волокон находитсяв прямой зависимости от активности миозин-АТФ-азы
- фермента, расщепляющего АТФ и тем самым
способствующего образованию поперечных мостиков
и взаимодействию актиновых и миозиновых
миофиламентов. Более высокая активность этого
фермента в быстрых мышечных волокнах
обеспечивает и более высокую скорость их
сокращения по сравнению с медленными волокнами
Тонус – слабое общее напряжение мышц
(развивается при очень низкой частоте стимуляции).
Сила и скорость сокращения мышц зависит от
количества вовлеченных в сокращение двигательных
единиц (чем больше двигательных единиц
активировано – тем сильнее сокращение).
Рефлекторный тонус - (наблюдается у некоторых
групп позных мышц) состояние непроизвольного
устойчивого напряжения мышц
33.
КПД мышцыВо время активации мышцы повышение
внутриклеточной концентрации Са 2+ ведет к
сокращению и к усиленному расщеплению АТФ; при
этом интенсивность метаболизма мышцы возрастает
в 100-1000 раз. Согласно первому началу
термодинамики (закону сохранения энергии),
химическая энергия, высвобождаемая в мышце,
должна быть равна сумме механической энергии
(мышечной работы) и теплообразования
34.
Коэффициент полезного действия.Гидролиз одного моля АТФ дает 48 кДж энергии,
40 –50% - превращается в механическую работу, а
50-60% рассеивается в виде тепла при запуске
(начальная теплота) и во время сокращения
мышцы, температура которой при этом
повышается. Однако в естественных условиях
механический КПД мышц около 20-30% так как во
время сокращения и после него процессы
требующие затрат энергии, идут и вне
миофибрилл (работа ионных насосов,
окислительная регенерация АТФ – теплота
восстановления)
35.
Энергетическийметаболизм
.
Во
время
продолжительной
равномерной
мышечной
активности происходит аэробная регенерация АТФ за
счет
окислительного
фосфорильирования.
Необходимая для этого энергия выделяется в
результате окисления углеводов и жиров. Система
находится в состоянии динамического равновесия –
скорости образования и расщепления АТФ равны.
(внутриклеточные
концентрации
АТФ
и
креатинфосфата относительно постоянны) При
продолжительных спортивных нагрузках скорость
расщепления АТФ в мышцах возрастает в 100 или в
1000 раз. Продолжительная нагрузка возможна если
скорость
восстановления
АТФ
возрастает
соответственно расходу. Потребление кислорода
мышечной тканью возрастает в 50-100 раз;
повышается скорость расщепления гликогена в
мышцах.
36.
Анаэробное расщепление – гликолиз: АТФ образуется в 2-3раза быстрее, а механическая энергия мышцы в 2-3 раза
выше, чем при длительной работе, обеспечиваемой
аэробными механизмами. Но ресурсы для анаэробного
метаболизма быстро исчерпываются, продукты метаболизма
(молочная кислота) вызывают метаболический ацидоз.,
который ограничивает работоспособность и вызывает
утомление. Анаэробные процессы необходимы для
обеспечения энергией кратковременного экстремального
усилия, а так же в начале продолжительной мышечной
работы, потому что адаптация скорости окисления (и
гликолиза) к возросшей нагрузке требует некоторого времени.
Кислородная задолженность приблизительно соответствует
количеству энергии, полученному анаэробным путем, еще не
компенсированное за счет аэробного синтеза АТФ.
Кислородная задолженность обусловлена(анаэробным)
гидролизом креатинфосфата, может достигать 4 л и может
увеличиваться до 20 л. Часть лактата окисляется в миокарде
а часть(преимущественно в печени) используется для синтеза
гликогена.
37.
Соотношение быстрых, и медленных волокон. Чембольше быстрых волокон содержит мышца, тем больше
возможная ее сила сокращения.
Поперечное сечение мышцы.
Термины «абсолютная» и «относительная» сила мышцы:
«общая сила мышцы» (определяется максимальным
напряжением в кг, которое она может развить) и «удельная
сила мышцы» - отношение этого напряжения в кг к
физиологическому поперечному сечению мышцы (кг/см2).
Чем больше физиологическое поперечное сечение мышцы,
тем больший груз она в состоянии поднять. По этой причине
сила мышцы с косо расположенными волокнами больше
силы, развиваемой мышцей той же толщины, но с
продольным расположением волокон. Для сравнения силы
разных мышц максимальный груз, который они в состоянии
поднять, делят на плошадь их физиологического поперечного
сечения (удельная сила мышцы). Вычисленная таким образом
сила (кг/см2) для трехглавой мышцы плеча человека - 16,8,
двуглавой мышцы плеча - 11,4, сгибателя плеча - 8,1,
икроножной мышцы - 5,9, гладких мышц - 1 кг/см2.
38.
В различных мышцах тела соотношение междучислом медленных и быстрых мышечных волокон
неодинаково, поэтому и сила их сокращения, и
степень укорочения вариабельны.
При снижении физической нагрузки - особенно
большой интенсивности, при которой требуется
активное участие быстрых мышечных волокон, последние истончаются (гипотрофируются) быстрее,
чем медленные волокна, быстрее уменьшается их
число
Факторы, влияющие на силу сокращения мышцы.
Число сокращающихся волокон в данной мышце. С
увеличением сокращающихся волокон возрастает
сила сокращений мышцы в целом. В естественных
условиях сила сокращения мышцы возрастает с
увеличением нервных импульсов, поступающих к
мышце,
в эксперименте - с увеличением силы раздражения.
39.
Умеренное растяжение мышцы также ведет кувеличению ее сократительного эффекта. Однако
при чрезмерном растяжении сила сокращения
уменьшается. Это демонстрируется в опыте с
дозированным растяжением мышцы: мышца
перерастянута так, что нити актина и миозина не
перекрываются, то общая сила мышцы равна нулю.
По мере приближения к натуральной длине покоя,
при которой все головки миозиновых нитей способны
контактировать с актиновыми нитями, сила
мышечного сокращения вырастает до максимума.
Однако при дальнейшем уменьшении длины
мышечных волокон из-за перекрытия нитей актина и
миозина сила сокращения мышцы снова
уменьшается вследствие уменьшения возможной
зоны контакта нитей актина и миозина.
40.
Функциональное состояние мышцы.При утомлении мышцы величина ее сокращения
снижается.
Работа мышцы измеряется произведением
поднятого груза на величину ее укорочения.
Зависимость мышечной работы от нагрузки
подчиняется закону средних нагрузок. Если мышца
сокращается без нагрузки, ее внешняя работа равна
нулю. По мере увеличения груза работа
увеличивается, достигая максимума при средних
нагрузках. Затем она постепенно уменьшается с
увеличением нагрузки. Работа становится равной
нулю при очень большом грузе, который мышца при
своем сокращении не способна поднять напряжение
100-200 мг.
41.
ГЛАДКАЯ МЫШЦА.Гладкая мускулатура не имеет поперечную
исчерченность. Клетки в виде веретен соединены
особыми межклеточными контактами (десмосомами).
Скорость скольжения миофибрилл и расщепления АТФ
ниже в 100-1000 раз. Хорошо приспособлены для
длительного устойчивого сокращения, которое не
приводит к утомлению и значительным энергозатратам.
Способны к спонтанным тетанообразным сокращениям,
которые имеют миогенное происхождение, а не
нейрогенное как у скелетных мышц.
Миогенное возбуждение.
Миогенное возбуждение возникает в клетках
ритмоводителях (пейсмекерах), которые обладают
электрофизиологическими свойствами.
Пейсмекерные потенциалы деполяризуют их мембрану
до порогового уровня, вызывая потенциал действия. Са
2+ поступает в клетку – мембрана деполяризуется, потом
42.
Спонтанную активность пейсмекеров можно модулироватьвегетативной нервной системой и ее медиаторами
(ацетилхолин усиливает активность приводя к более частым и
сильным сокращениям, а норадреналин оказывает
противоположное действие).
Возбуждение распространяется через «щелевые контакты»
(нексусы) между плазматическими мембранами
сопредельных мышечных клеток. Мышца ведет себя как
единая функциональная единица, синхронно воспроизводя
активность своего пейсмекера. Гладкая мышца может быть
полностью расслаблена как в укороченном так и в растянутом
состоянии. Сильное растяжение активирует сокращение.
Электромеханические сопряжение. Возбуждение
гладкомышечных клеток вызывает либо увеличение входа Са
через потенциалзависимые кальциевые каналы, либо
высвобождает из кальциевых депо, что в любом случае
приводит к возрастанию внутриклеточной концентрации
кальция и вызывает активацию сократительных структур.
Расслабление идет медленно т.к. скорость поглощения ионов
Са очень низкая.


