Скелетная мышечная ткань. Шпаргалка: Мышечные ткани
Тема: «Мышечные ткани»
Вопрос 1 . Общие структурные особенности мышечных тканей
Объединяет несколько разных видов, но основное свойство общее – сократимость. Поэтому все мышечные ткани имеют сходные структурные особенности:
Клетки вытянутой формы и объединены в тяжи, или даже в симпласты (мышечные волокна).
Цитоплазма заполнена миофиламентами – нитями из сократительных белков (миозин и актин), взаимное скольжение которых обеспечивает сокращение. Характер расположения миофиламентов зависит от вида мышечной ткани.
Высокие энергетические запросы требуют множества митохондрий, включений миоглобина, жира и гликогена.
Гладкая ЭПС специализирована на накоплении Сa2+, который иницииирует сокращение.
Плазмолемма мышечных клеток обладает возбудимостью.
Согласно морфо-функциональной классификации выделяют:
Поперечно-полосатые мышечные ткани. В их цитоплазме главный компонент – миофибриллы (органеллы общего значения), который и создают эффект исчерченности. Этих тканей два вида:
скелетная. Образуется из миотомов сомитов.
сердечная. Образуется из висцерального листка спланхнотома.
Гладкая мышечная ткань. Ее клетки не содержат миофибрилл. Образуется из мезенхимы.
К этой же группе относят миоэпителиальные клетки, которые имеют эктодермальное происхождение и мышцы радужки глаза, которые имеют нейральное происхождение.
Вопрос 2 . Скелетная мышечная ткань Организация мышечного волокна
Структурно-функциональной единицей этой ткани является мышечное волокно. Это длинный цитоплазматический тяж со множеством ядер, которые лежат сразу под плазмолеммой. Мышечное волокно в эмбриогенезе образуется при слиянии клеток – миобластов, т.е., представляет собой клеточное производное –симпласт.
Мышечное волокно сохраняет общий план клеточной организации. В нем есть все органеллы общего значения, много включений, а также органеллы специального значения. Все компоненты волокна адаптированы для выполнения главной функции – сокращения – и подразделяются на несколько аппаратов.
Сократительный аппарат состоит из миофибрилл. Это органеллы, которые тянутся вдоль всего волокна и занимают большую часть всего объема цитоплазмы. Они способны значительно изменять свою длину.
Аппарат белкового синтеза представлен, в основном, свободными рибосомами и специализирован на выработке белков для построения миофибрилл.
Аппарат передачи возбуждения образован саркотубулярной системой. Она включает гладкую ЭПС и Т-трубочки. Гладкая ЭПС (саркоплазматическая сеть) имеет вид плоских цистерн, которые оплетают все миофибриллы. Она служит для накопления Сa2+. Ее мембраны способны быстро выпускать кальций наружу, что необходимо для укорочения миофибрилл, а затем активно закачивает его внутрь. Наружная мембрана мышечного волокна (сарколемма) образует многочисленные трубчатые впячивания, пронизывающие все волокно в поперечных направлениях. Их совокупность называют Т-системой. Т-трубочки тесно контактируют с мембранами ЭПС, образуя единую саркотубулярную систему. К каждой Т-трубочке …..
Энергетический аппарат составлен митохондриями и включениями. Митохондрии крупные вытянутые и лежат, в основном цепочками, заполняя все пространство между миофибриллами. Субстратами для получения АТФ служит гликоген и липидные капли. Включения миоглобина – специфического мышечного пигмента, обеспечивают волокна кислородом в случае длительной и напряженной работы мышц.
Лизосомальный аппарат развит слабо. Служит, главным образом, для процессов внутриклеточной регенерации.
Вопрос 3 Механизм мышечного сокращения
Для его понимания необходимо ознакомиться с молекулярной организацией миофибрилл – органелл, специализированных на сокращении. Это длинные тяжи, образующие продольные пучки по тысяче и более миофибрилл, которые почти полностью заполняют цитоплазму волокна.
Каждая миофибрилла построена из огромного числа актиновых и миозиновых филаментов.
Тонкие актиновые нити построены из глобулярных молекул белка актина, которые объединяются в две спирально закрученные цепочки. Более толстая миозиновая нить построена из 300-400 молекул белка миозина. Каждая молекула включает длинный хвост, к которому с одного края прикреплена подвижная головка. Головки могут менять угол своего наклона. Хвосты множества молекул укладываются плотным пучком, формируя стержень филамента. Головки при этом остаются на поверхности. На двух краях нити головки лежат разнонаправленно.
Благодаря дополнительным белкам, миофиламенты имеют стабильный диаметр и стабильную длину около 1 мкм. Филаменты одного вида образуют аккуратно подогнанные пучки или стопки. Миофибриллы образованы из многократно чередующихся пучков актиновых и миозиновых нитей. Высокая упорядоченность в расположении миофиламентов достигается с помощью различных белков цитоскелета. Например, белок актинин формирует Z-линию (телофрагму), к которой с обеих сторон присоединяются края тонких актиновых нитей. Так образуется актиновая стопка. Другие белки организуют в стопку толстые миозиновые нити, прошнуровывая их в середине. Они образуют М-линию (мезофрагму). При чередовании стопок свободные концы тонких и толстых нитей заходят друг за друга, обеспечивая взаимное скольжение друг относительно друга в момент сокращения. В результате такой организации в миофибрилле многократно повторяются светлые участки, называемые I-дисками (изотропные), и темные участки, называемые А-дисками (анизотропные). Это и создает эффект поперечной исчерченности. Изотропные участки соответствуют центральной части актиновой стопки и содержат только тонкие нити. Анизотропные диски соответствуют целой миозиновой стопке, и в них входят чисто миозиновая часть (Н-полоска) и те участки, где концы тонких и толстых нитей перекрываются.
Участок между двумя Z-линиями называют саркомером. Саркомер является структурной единицей миофибриллы. (20 тысяч саркомеров на миофибриллу). Строгая организация миофибрилл обеспечивается широким набором различных белков цитоскелета.
При сокращении длина миофибриллы уменьшается за счет одновременного укорочения всех I-дисков. Длина каждого саркомера при этом уменьшается в 1,5-2 раза. Процесс сокращения объясняется теорией скольжения Хаксли, согласно которой в момент сокращения свободные, заходящие друг за друга концы актиновых и миозиновых нитей вступают в молекулярные взаимодействия и глубже задвигаются друг относительно друга. Скольжение начинается с того, что выступающие миозиновые головки образуют мостики с активными центрами актиновых филаментов. Затем угол наклона головки уменьшается, мостики совершают как бы гребковые движения, смещая и актиновую нить. После этого мостик размыкается, что сопровождается гидролизом 1 молекулы АТФ. Связывание миозиновых головок с актиновой нитью регулируется специальными белками. Это тропонин и тропомиозин, которые встроены в актиновую нить, и препятствуют контакту с миозиновыми головками. При повышении в гиалоплазме концентрации Са2+ происходит изменение конформационного состояния этих регуляторных белков и их блокирующее действие снимается. Гребковые движения повторяются сотни раз за одно мышечное сокращение. Расслабление наступает только после того, как снизится концентрация Ca2+.
Вопрос 4. Аппарат передачи возбуждения
Сокращение запускается нервным импульсом, который через моторную бляшку передается на мембрану мышечного волокна, вызывая волну деполяризации, которая мгновенно охватывает и Т-трубочки. Они тянутся от поверхности сквозь все волокно, по пути колечками окружая миофибриллы. Полости гладкой ЭПС, заполненные кальцием, чехлом оплетают миофибриллы, тесно контактируя с Т-трубочками. С двух сторон к каждой Т-трубочке прилежат обширные мембранные полости ЭПС (терминальные цистерны). Такой комплекс называют триадой. На каждый саркомер приходится две триады. Благодаря мембранным контактам деполяризация Т-трубочек изменяет состояние мембранных белков ЭПС, что приводит к открытию кальциевых каналов и выходу кальция в гиалоплазму. Происходит сокращение. Триады сопрягают процессы возбуждения и сокращения. После выброса специальные мембранные насосы активно закачивают Ca2+ обратно в ЭПС, где он соединяется с Са-связывающим белком.
Вопрос 5. Сердечная мышечная ткань
образует мышечную стенку сердца – миокард. Ее морфо-функциональная единица – отдельная клетка – кардиомиоцит. Клетки соединены друг с другом особыми структурами – вставочными дисками, и в результате образуется трехмерная сеть из клеточных тяжей (функциональный синцитий), что обеспечивает синхронность сокращения во время систолы.
Кардиомиоциты – вытянутые клетки с несколькими ответвлениями, покрытые поверх плазмолеммы базальной мембраной. Их ядра (1 или 2) лежат центрально.
В составе миокарда выделяют несколько популяций кардиомиоцитов:
А) сократительные или рабочие
Б) проводящие
В) секреторные
Вопрос 6. Рабочие кардиомиоциты
составляют основную массу миокарда и обеспечивают сокращение. Их организация сходна с мышечными волокнами, но имеет ряд отличий.
сократительный аппарат. Миофибриллы в целом имеют продольное направление, но многократно анастомозируют друг с другом.
Саркотубулярная сеть развита слабее. Т-трубочки более широкие, лежат реже и каждая контактирует только с одной цистерной ЭПС (диада). При возбуждении часть Ca2+ поступает в гиалоплазму из межклеточного пространство через плазмолемму и мембраны Т-трубочек и лишь после этого происходит Са-индуцированный выброс Ca2+ из ЭПС.
Энергетический аппарат. Митохондрии много, они крупные с плотно упакованными кристами, поскольку энергетические запросы миокарда очень высоки. Между собой они объединены особыми соединениями – межмитохондриальными контактами и образуют единую функциональную систему – митохондрион. Такая интеграция исключительно важна для быстрого и синхронного сокращения миокарда. Субстраты для получения АТФ поставляются липидными каплями, включениями гликогена и миоглобина. Сами мотохондрии способны накапливать кальций.
Концы соседних клеток или их стыкующиеся ответвления соединяются вставочными дисками. Диск имеет ступенчатую форму. Поперечные участки образованы десмосомами и придают соединению механическую прочность. Продольные участки содержат множество щелевых контактов – нексусов, которых особенно много в предсердиях. Благодаря ионным каналам нексусов возбуждение быстро распространяется вдоль всей мышцы.
Миокард обильно кровоснабжается. Все промежутки между кардиомиоцитами заполнены рыхлой соединительной тканью, в которой ветвятся капилляры. Здесь же заканчиваются ветвления нервных волокон вегетативной нервной системы. В отличие от скелетной мышечной ткани здесь образуются не нейро-мышечные синапсы (моторные бляшки), а лишь варикозные расширения. На сократительную активность кардиомиоцитов нервная система оказывает лишь регуляторной влияние. Вегетативная система лишь увеличивает (симпатический отдел) или уменьшает (парасимпатический отдел) частоту и силу сердечных сокращений.
PAGE_BREAK--
Ритмичная генерация импульсов, которые заставляют сердце постоянно сокращаться, обеспечивается специальными клетками самого миокарда. Совокупность этих клеток называется проводящей системой сердца, а способность сердца сокращаться независимо от нервных стимулов – автоматией сердца.
Вопрос 7 . Проводящая система
включает специализированные кардиоммиоциты, называемые также атипичными. К ним относят:
пейсмекерные клетки или водители ритма. Их главное свойство – неустойчивые потенциал покоя наружной мембраны. Благодаря К/Na -насосу натрия всегда больше внутри клетки, а калия снаружи. Эта разность ионов и создает электрический потенциал по обе стороны плазмолеммы. При определенной стимуляции в мембране открываются натриевые каналы, натрий устремляется наружу и мембрана деполяризуется. У пейсмекерных клеток благодаря постоянной небольшой утечке ионов плазмолемма регулярно деполяризуется без всяких внешних сигналов. Это вызывает потенциал действия, распространяющийся и на соседние клетки, вызывая их сокращение. Главные водители ритма – это кардиомиоциты синусно-предсердного узла. Каждую минуту они генерируют 60-90 импульсов. Водители ритма второго порядка образуют предсердно-желудочковый узел. Они генерируют импульсы с частотой 40 импульсов в минуту, и в норме их активность подавляется главными пейсмекерами. Пейсмекерные кардиомиоциты – мелкие светлые клетки с крупным ядром. Их сократительный аппарат развит слабо.
Проводящие кардиомиоциты обеспечивают быструю передачу возбуждения от водителей ритма к рабочим кардиомиоцитам. Эти клетки объединены в длинные тяжи, формирующие пучок Гиса и волокна Пуркинье. Пучок Гиса составлен клетками среднего размера с редкими длинными извилистыми миофибриллами и мелкими митохондриями. Волокна Пуркинье содержат самые крупные кардиомиоциты, которые могут контактировать сразу с несколькими рабочими клетками. Миофибриллы здесь образуют редкую неупорядоченную сеть, Т-система не развита. Вставовных дисков нет, но клетки объединены множеством нексусов, что обеспечивает высокую скорость проведения импульсов.
Вопрос 8. Секреторные кардиомиоциты
В предсердиях встречаются отросчатые клетки, в которых хорошо развита грЭПС, комплекс Гольджи и содержатся секреторные гранулы. Миофибриллы развиты очень слабо, поскольку основной функцией является выработка гормона (натрийуретический фактор), регулирующего обмен электролитов и артериальное давление.
Вопрос 9 . Гладкая мышечная ткань
Построена из гладких миоцитов. Сократительные филаменты в этих клетках не имеют жесткой упорядоченности и миофибриллы в них не образуются. Вследствие этого отсутствует и поперечная исчерченность. Гладкие миоциты довольно крупные клетки веретеновидной формы, покрытые сверху базальной мембраной, которая соединена с межклеточным веществом. В центре вытянутое ядро, у полюсов грЭПС, комплекс Гольджи и рибосомы. Клетки секретируют компоненты межклеточного вещества для своей наружной оболочки, а также некоторые ростовые факторы и цитокины. Много мелких митохондрий. Саркоплазматическая сеть (гладкая ЭПС) развита слабо, она выполняет роль кальциевого депо. Системы Т-трубочек нет, и их функцию выполняют кавеолы. Кавеолы – это мелкие впячивания плазмолеммы в виде пузырьков. Они содержат высокие концентрации кальция, который захватывают из межклеточного пространства. В момент возбуждения Ca2+ из кавеол выходит наружу, что инициирует освобождение Ca2+ из саркоплазматической сети.
Организация и функционирование сократительного аппарата своеобразны. Актиновые и миозиновые филамента очень многочисленны, но не образуют миофибрилл. Для их упорядочивания в миоците существует система плотных телец. Это округлые опорные образования из белка a-актинина и десмина. В них одним концом закреплено по 10-20 тонких актиновых филаментов. Одни тельца образуют прикрепительные пластинки в сарколемме, другие цепочками лежат прямо в гиалоплазме. Так в миоците формируется стабильная сеть из актиновых нитей. Толстые миозиновын нити имеют разную длину и очень лабильны.
Каждому сокращению предшествует выброс кальция, который связывается с особым белком – кальмодулином. Это активирует фермент, обеспечивающий быструю сборку миозиновых филаментов. Они встраиваются между актиновыми нитями, образуют с ними мостики, и их головки начинают совершать гребковые движения. При взаимном скольжении нитей плотные тельца сближаются, а клетка в целом укорачивается. Таким образом в гладких миоцитах кальций взаимодействует с миозиновыми нитями, а не с актиновыми, как в поперечно-полосатых. АТФ-азная активность миозина намного ниже. Вместе с постоянной сборкой и разборкой миозиновых филаментов это приводит к тому, что гладкомышечные клетки сокращаются медленнее, но могут длительно поддерживать этот состояние (тонические сокращения). Между собой клетки объединены рвст, которая вплетается в их базальные мембраны, а также различными межклеточными контактами, в том числе и нексусами. Сократительная активность миоцитов находится под контролем нервных и гуморальных факторов. В соединительно-тканных прослойках расположены варикозные расширения аксонов вегетативной нервной системы. Их медиаторы деполяризуют ближайшие миоциты, а к остальным возбуждение передается по щелевидным контактам.
Благодаря широкому набору мембранных рецепторов гладкие миоциты чувствительны ко многим биологически активным веществам (адреналин, гистамин и т.д.) и реагируют по разному, в зависимости от органной специфичности.
Вопрос 10. Гистогенез и регенерация
Скелетная мышечная ткань. Из миотома сомитов дифференцируются одноядерные активно делящиеся клетки – миобласты. Они сливаются в цепочки - мышечные трубочки, многочисленные ядра которых уже не делятся. В трубочках начинается активный синтез сократительных белков и формирование миофибрилл, которые постепенно заполняют всю цитоплазму, оттесняя ядра на периферию. Образуется мышечное волокно, внутри которго миофибриллы постоянно обновляются. Между плазмолеммой и покрывающей ее базальной мембраной кое-где сохраняются одноядерные клетки – миосаттелиты – камбиальные клетки, которые могут делиться и включать свои ядра в состав волокон. Рост мышечной ткани у взрослого человека происходит, в основном за счет гипертрофии волокон, а их число остается постоянным. После повреждения миосаттелиты могут сливаться, образуя новые волокна.
Сердечная мышечная ткань образуется из миоэпикардиальной пластинки в составе висцерального листка спланхнотома. Деление кардиомиоцитов заканчивается вскоре после рождения, но в последующие 10 лет могут формироваться полиплоидные и двуядерные клетки. Поскольку камбиальных клеток нет, то возможна только внутриклеточная регенерация и гипертрофия кардиомиоцитов. Она происходит в результате длительных физических нагрузок, либо в патологических состояниях (гипертония, пороки сердца и т.д.). После гибели миоцитов (инфркт миокарда) формируется соединительно-тканный рубец. В последнее время установлено, что отдельные предсердные миоциты сохраняют способность к митозам.
Гладкомышечная ткань регенерирует как за счет гипертрофии, так и за счет гиперплазии
Мышечными тканями называют ткани, различные по строению и происхождению, но сходные по способности к выраженным сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма.
Источник развития: поперечнополосатая скелетная мышечная ткань относится к соматическому типу, развивается из миотомов сомитов, образует скелетную поперечнополосатую мускулатуру.
Основной структурной единицей скелетной мышечной ткани является поперечнополосатое мышечное волокно, состоящее из миосимпласта и миосателитоцитов, покрытых общей базальной мембраной.
Мышечное волокно – цилиндрическое образование, диаметром от 10 до 100 мкм и длиной до 12 см. Мышечные волокна в мышцах образуют пучки, в кот. они лежат параллельно.
Миосателитоциты имеют узкий узкий ободок цитоплазмы и удлиненное ядро с крупными глыбками хроматина, слабое развитие органелл. Расположены эти клетки между плазмолеммой миосимпласта и базальной мембраной. Являются главным источником развития мышечных волокон в процессе посттравматической регенерации мышц.
Миосимпласт содержит от нескольких сотен тысяч ядер, лежащих на периферии под плазмолеммой. Ядра светлые, овальные, диплоидные, уплощенные, длиной 10-20 мкм, располагаются на расстоянии 5 мкм друг от друга. Саркоплазма миосимпласта содержит все органеллы общего значения(кроме центриолей) и специальные органеллы и включения. Эти структуры образуют функциональные аппараты:
- Сократительный аппарат миосимпласта представлен миофибриллами, кот. имеют вид нитей диаметром 1-2мкм и расположены в центральной части саркоплазмы и отделены друг от друга митохондриями. Миофибриллы обладают собственной поперечной исчерченностью, кот. обусловлена чередованием темных А-дисков (анизотропных, обладающих двойным лучепреломлением) и светлых И-дисков (изотропных, не обладающих двойным лучепреломленим).
В середине И-диска – телофрагма, а в середине А-диска проходит М-линия, образованная утолщением миозиновых нитей.
Саркомер - структурная единица миофибриллы. Саркомер – участок миофибриллы между двумя телофрагмами, состоит из А-диска, расположенного в центре, и двух половинок И-диска, расположенных на периферии саркомера. В расслабленной мышце длина саркомера 2-3 мкм, при сокращении- он укорачивается до 1,5мкм.
Саркомер- система толстых и тонких белковых нитей- миофиламентов. Толстые нити образованы молекулами белка миозина и расположены в А-диске, а тонкие нити содержат сократимый белок актин и два регуляторных белка – тропин и тропомиозин, тонкие нити прикрепляются к телофрагмам, образуют И-диски и частично проникают в А-диски между толстыми нитями. Участок А-диска, свободный от тонких волокон, называется светлой полоской Н.
Формула саркомера: Т+ И/2 +А/2 + Н/2 + М + Н/2 + А/2 + И/2 + Т
- Трофический аппарат – вся масса саркоплазмы вместе с органеллами и ядрами. Много митохондрий между миофибриллами, по периферии саркомеров у телофрагмы
- Специфический мембранный аппарат состоит из: 1) саркоплазматическая сеть – состоит из утолщенных каналов и терминальных цистерн, в кот.происходит накопление ионов кальция. Каналы расположены вдоль саркомеров между миофибриллами и названы Л-каналами. Цистерны расположены вблизи телофрагм. 2) канальцы Т-системы – впячивания плазмолеммы поперек оси волокна и локализуются на уровне телофрагм. Этот аппарат образует триады (один каналец в центре и две цистерны по краям), сюда поступает нервный импульс, кот. распространяется вглубь мышечного волокна.
- Опорный аппарат представлен а) внешним компонентом – сарколеммой - это плазмолемма, снаружи кот. прилежит базальная мембрана, в кот. вплетены ретикулярные и коллагеновые волокна и б) внутренним компонентом – телофрагмы, кот. вплетаются в плазмолемму и укрепляют волокно в поперечном направлении.
- Щитовидная железа: источники развития. Строение структурно-функциональной единицы при нормо- гипо- и гиперфункции. Роль железы. Регуляция ее деятельности.
Щитовидная железа – содержит 2 типа эндокринных клеток: фолликулярные эндокриноциты, тироциты (вырабатывают тироксин), и парафолликулярные эндокриноциты (вырабатывают кальцитонин).
Источник развития : зачаток ЩЖ возникает у зародыша человека на 3-4 неделе как выпячивание стенки глотки между I и II парами жаберных карманов, кот. растет вдоль глоточной кишки в виде эпителиальноо тяжа. На уровне III-IV пар жаберных карманов этот тяж раздваивается, давая начало формирующимся правой и левой долям ЩЖ. Зачатки долей быстро разрастаются, образуя рыхлые сети ветвящихся эпителиальных трабекул, из них формируются тироциты, образующие фолликулы, в промежутки между кот. врастает мезенхима с кров.сосудами и нервами.
Строение ЩЖ:
Железа окружена соединительной капсулой, прослойки которой разделяют орган на дольки. Основными структурными компонентами паренхимы являются – фолликулы – замкнутые шаровидные или слегка вытянутые образования с полостью внутри, образованные одним слоем эпителиальных клеток, представленных фолликулярными эндокриноцитами (тироцитами), а также парафолликулярными эндокриноцитами нейрального происхождения. В просвете фолликулов накапливается коллоид – секреторный продукт тироцитов – вязкая жидкость, состоящая из тироглобулина. Диаметр фолликула от 0,02 до 0,9 мм. В фолликулах, не заполненных коллоидом, эпителий призматический. По мере накопления коллоида размер фолликулов увеличивается, эпителий становится кубическим, а в сильно растянутых фолликулах – плоским.
Фолликулы разделяются прослойками РВСТ с кровеносными и лимфатическими капиллярами, тучными клетками и лимфоцитами.
Фолликулярные эндокриноциты (тироциты) – железистые клетки, составляют большую часть стенки фолликулов. Тироциты располагаются в один слой на базальной мембране. При умеренной функциональной активности ЩЖ тироциты имеют кубическую форму и шаровидные ядра. Коллоид заполняет просвет фолликула. На апикальной поверхности тироцитов есть микроворсинки. По мере усиления тироидной активности количество и размеры микроворсинок возрастают. Одновременно базальная поверхность тироцитов, почти гладкая в период функционального покоя ЩЖ, становится складчатой. По мере возрастания тироидной активности на боковых поверхностях тироцитов возникают пальцевидные выступы (интердигитиции), входящие в соответствующие вдавления боковой поверхности соседних клеток.
Белковые продукты, синтезируемые тироцитами, выделяются в полость фолликула, где завершается образование йодированных тирозинов и тиронинов (аминокислот).
Когда активность ЩЖ увеличивается (гиперфункция), тироциты принимают призматическую форму.Коллоид становится более жидким. Ослабление активности ЩЖ (гипофункция) – уплотнение коллоида, высота тироцитов уменьшается, принимают уплощенную форму.
Парафолликулярные эндокриноциты (кальцитониноциты) – в стенке фолликулов, в межфолликулярных прослойках соединительной ткани. Клетки не поглощают йод, но совмещают образование нейроаминов (норадреналин и серотонин) с синтезом белковых гормонов (кальцитонина и соматостатина).
Роль железы: ЩЖ содержит фолликулярные эндокриноциты,/тироциты (вырабатывают тироксин), и парафолликулярные эндокриноциты (вырабатывают кальцитонин).
Кальцитонин и паратгормон тесно взаимодействуют в регуляции минерально обмена. КАльцитонин снижает уровень кальция в крови и ингибирует образование кости остеобластами. Паратгормон повышает уровень кальция в сыворотке крови и стимулирует резорбцию кости остеокластами. Гипокальциемия усиливает секрецию паратгормона, а гиперкальциемия- подавляет. Кальцитонин и паратгормон действуют на функцию почек и гастроинтестинальный тракт, регулируя экскрецию и поглощение кальция в этих органах.
3. Морфо-функциональная характеристика кожного покрова. Источники развития. Потовые, сальные железы, их строение, функции.
Кожа образует внешний покров организма, площадь которого у взрослого человека достигает 2,5 м2. Кожа состоит из эпидермиса (эпителиальная ткань) и дермы (соединительнотканная основа). С подлежащими частями организма кожа соединяется слоем жировой ткани - подкожной клетчаткой, или гиподермой.
- Эпидермис представлен многослойным плоским ороговевающим эпителием, в котором постоянно происходят обновление и специфическая дифференцировка клеток (кератинизация).
На ладонях и подошвах эпидермис состоит из многих десятков слоев клеток, которые объединены в 5 основных слоев: базальный, шиповатый, зернистый, блестящий и роговой. В остальных участках кожи 4 слоя (отсутствует блестящий слой). В них различают 5 типов клеток: кератиноциты (эпителиоциты), клетки Лангерганса (внутриэпидермальные макрофаги), лимфоциты, меланоциты, клетки Меркеля. Из этих клеток в эпидермисе и каждом из его слое основу составляют кератиноциты. Они непосредственно участвуют в ороговении (кератинизации) эпидермиса.
Собственно кожа, или дерма , делится на два слоя - сосочковый и сетчатый, которые не имеют между собой четкой границы.
Функции кожи:
- Защитная – кожа защищает ткани от механических, химических и др. воздействий. Роговой слой эпидермиса препятствует проникновению в кожу микроорганизмов. Кожа принимает участие в обеспечении норм. водного баланса. Роговой слой эпидермиса обеспечивает преграду испаряющейся жидкости, предотвращает набухание и сморщивание кожи.
- Выделительная – вместе с потом через кожу в сутки выделяются около 500 мл воды, различные соли, молочная кислота, продукты азотистого обмена.
- Участие в терморегуляции – благодаря наличию терморецепторов, потовых желез и густой сети кров. сосудов.
- Кожа – депо крови. Сосуды дермы при их расширении могут вместить до 1 л крови
- Участие в обмене витаминов – под действием УФЛ в кератиноцитах синтезируется вит.D
- Участие в метаболизме многих гормонов, ядов, канцерогенов.
- Участие в иммунных процессах – в коже происходит распознавание антигенов и их элиминация; антигензависимая пролиферация и дифференцировка Т-лимфоцитов, иммунологический надзор над опухолевыми клетками(при участии цитокинов).
- Явл-ся обширным рецепторным полем, позволяющим ЦНС получать информациюоб изменении в самой коже и о характере раздражителя.
Источники развития . Кожа развивается из двух эмбриональных зачатков. Эпителиальный покров (эпидермис) ее образуется из кожной эктодермы, а подлежащие соединительнотканные слои - из дерматомов (производных сомитов). В первые недели развития зародыша эпителий кожи состоит всего из одного слоя плоских клеток. Постепенно эти клетки становятся все более высокими. В конце 2-го месяца над ними появляется второй слой клеток, а на 3-м месяце эпителий становится многослойным. Одновременно в наружных его слоях (в первую очередь на ладонях и подошвах) начинаются процессы ороговения. На 3-м месяце внутриутробного периода в коже закладываются эпителиальные зачатки волос, желез и ногтей. В соединительнотканной основе кожи в этот период начинают образовываться волокна и густая сеть кровеносных сосудов. В глубоких слоях этой сети местами появляются очаги кроветворения. Лишь на 5-м месяце внутриутробного развития образование кровяных элементов в них прекращается и на их месте формируется жировая ткань.
Железы кожи . В коже человека находятся три вида желез: молочные, потовые и сальные.
Потовые железы подразделяются на эккриновые (мерокриновые) и апокриновые.
Потовые железы по своему строению являются простыми трубчатыми. Они состоят из выводного протока и концевого отдела. Концевые отделы располагаются в глубоких частях сетчатого слоя на границе его с подкожной клетчаткой, а выводные протоки эккриновых желез открываются на поверхности кожи потовой порой. Выводные протоки многих апокриновых желез не заходят в эпидермис и не образуют потовых пор, а впадают вместе с выводными протоками сальных желез в волосяные воронки.
Концевые отделы эккриновых потовых желез выстланы железистым эпителием, клетки которого бывают кубической или цилиндрической формы. Среди них различают светлые и темные секреторные клетки.
Концевые отделы апокриновых желез состоят из секреторных и миоэпителиальных клеток. Переход концевого отдела в выводной проток совершается резко. Стенка выводного протока состоит из двухслойного кубического эпителия.
- участвуют в терморегуляции и способствуют выведению вместе с потом воды (500мл/сутки) и продуктов метаболизма веществ (мочевина, аммиак, мочевая кислота и др.)
Сальные железы являются простыми альвеолярными с разветвленными концевыми отделами. Секретируют они по голокриновому типу.
Концевые отделы представлены альвеолами или дольками, кот. состоят из клеток-себоцитов, среди которых различают недифференцированные, дифференцированные и некротические (погибающие) формы. Жедезы трех типов: 1) мелкие –кожа бровей,век; 2) средние 3) крупные – на лице и верхней части туловища.
Альвеола состоит из периферических клеток, дифференцированных клеток, зоелых клеток.
Выводной проток короткий. Стенка его состоит из многослойного плоского ороговевающего эпителия. Ближе к концевому отделу количество слоев в стенке протока уменьшается, эпителий становится кубическим и переходит в наружный ростковый слой концевого отдела.
У мужчин сальные железы секретируют до 80 лет, у женщин до 50 лет. Эстрогены угнетают деятельность сальных желез, прогестерон- наоборот. Тироидные гормоны активируют 5-альфа-редуктазу, усиливают деятельность себатоцитов.
- выполняют защитную функцию – предохраняют кожу от вредного действия химических веществ. Предотвращают высыхание кожи, участвуют в формировании защитного барьера на поверхности кожи.
- Характеристика дробления зародыша человека: тип дробления, продолжительность. Строение зародыша после дробления.
Дробление зародыша происходит в течение 4 суток после образования зиготы. Оно происходит в маточных трубах по мере продвижения зиготы в полость матки. Этот период называют трубным периодом эмбриогенеза. Яйцеклетка человека – вторично изолецитального типа. Дробление зародыша человека полное, неравномерное, асинхронное. С самого начала появляется полоса дробления по меридиану с образованием 2 бластомеров неодинаковой величины. Затем образуется 3 бластомера. Увеличение числа бластомеров происходит в начале медленно. Через 30 часов образуются 2 бластомера, через 40 часов – 4 бластомера. В ходе дробления появляются крупные темные бластомеры и мелкие светлые бластомеры. В процессе дробления мелкие бластомеры располагаются снаружи от крупных. Мелкие называются трофобластами, а крупные – эмбриобластами. Трофобласты обеспечивают трофику эмбриобласта.
К 3 суткам дробление ускоряется и к середине 3 суток образуется зародыш, кот. называется морулой, в ней отсутствует полость.
В начале 4 суток формируется бластоцист, имеющий полость. Этот зародыш находится в маточной трубу и к 5 суткам состоит из 58 бластомеров.
К концу 5- началу 6 суток бластоцист попадает в полость матки. К этому времени он состоит из 108 бластомеров.
К концу 7 суток зародыш находится в матке в свободном виде и подготавливается к имплантации – внедрению в эндометрий.
На стадии свободной бластоцисты (7-7,5суток) происходит первая фаза гаструляции путем миграции и деляминации – расслоение зародышевого диска. Образуется два зародышевых листка- эпибласт и гипобласт.
После их образования начинается имплантация зародыша в эндометрий матки.
Подавляющая часть гладкой мышечной ткани организма (внутренних органов и сосудов) имеет мезенхимальное происхождение.
Структурно-функциональной единицей гладкой мышечной ткани внутренних органов и сосудов является миоцит . Представляет собой чаще всего веретенообразную клетку (длиной 20-500 мкм, диаметром 5-8 мкм), покрытую снаружи базальной пластинкой, но встречаются и отростчатые миоциты. В центре располагается вытянутое ядро, по полюсам которого локализуются общие органеллы: зернистая эндоплазматическая сеть, пластинчатый комплекс, митохондрии, цитоцентр. В цитоплазме содержатся толстые (17 нм) миозиновые и тонкие (7 нм) актиновые миофиламенты, которые располагаются в основном параллельно друг другу вдоль оси миоцита и не образуют А и I диски, чем и объясняется отсутствие поперечной исчерченности миоцитов. В цитоплазме миоцитов и на внутренней поверхности плазмолеммы встречаются многочисленные плотные тельца, к которым прикрепляются актиновые, миозиновые, а так же промежуточные филаменты. Плазмолемма образует небольшие углубления - кавеолы, которые рассматриваются как аналоги Т-канальцев. Под плазмолеммой локализуются многочисленные везикулы, которые вместе с тонкими канальцами цитоплазмы являются элементами саркоплазматической сети.
Механизм сокращения в миоцитах в принципе сходен с сокращением саркомеров в миофибриллах в скелетных мышечных волокнах. Он осуществляется за счет взаимодействия и скольжения актиновых миофиламентов вдоль миозиновых. Для такого взаимодействия также необходимы энергия в виде АТФ, ионы кальция и наличие биопотенциала. Биопотенциалы поступают от эфферентных окончаний вегетативных нервных волокон непосредственно на миоциты или опосредованно от соседних клеток через щелевидные контакты и передаются через кавеолы на элементы саркоплазматической сети, обуславливая выход из них ионов кальция в саркоплазму. Под влиянием ионов кальция развиваются механизмы взаимодействия между актиновыми и миозиновыми филаментами, аналогичные тем, которые происходят в саркомерах скелетных мышечных волокон, в результате чего происходит скольжение названных миофиламентов и перемещение плотных телец в цитоплазме. В миоцитах, кроме актиновых и миозиновых филаментов, имеются еще промежуточные, которые одним концом прикрепляются к цитоплазматическим плотным тельцам, а другим - прикрепительным тельцам на плазмолемме и таким образом передают усилия взаимодействия актиновых и миозиновых филаментов на сарколемму миоцита, чем и достигается его укорочение.
Миоциты окружены снаружи рыхлой волокнистой соединительной тканью - эндомизием и связаны друг с другом боковыми поверхностями. При этом, в области тесного контакта соседних миоцитов базальные пластинки прерываются. Миоциты соприкасаются непосредственно плазмолеммами и в этих местах имеются щелевидные контакты, через которые осуществляется ионная связь и передача биопотенциала с одного миоцита на другой, что приводит к одновременному и содружественному их сокращению. Цепь миоцитов, объединенных механической и метаболической связью, составляет функциональное мышечное волокно. В эндомизии проходят кровеносные капилляры, обеспечивающие трофику миоцитов, а в прослойках соединительной ткани между пучками и слоями миоцитов в перимизии проходят более крупные сосуды и нервы, а также сосудистые и нервные сплетения.
Эфферентная иннервация гладкой мышечной ткани осуществляется вегетативной нервной системой. При этом, терминальные веточки аксонов эфферентных вегетативных нейронов, проходя по поверхности нескольких миоцитов, образуют на них небольшие варикозные утолщения, которые несколько прогибают плазмолемму и образуют мионевральные синапсы . При поступлении нервных импульсов в синаптическую щель выделяются медиаторы (ацетилхолин или норадреналин), и обуславливают деполяризацию мембран миоцитов и последующее их сокращение. Через щелевидные контакты биопотенциалы переходят из одного миоцита на другой, что сопровождается возбуждением и сокращением и тех гладкомышечных клеток, которые не содержат нервных окончаний. Возбуждение и сокращение миоцитов обычно продолжительны и обеспечивают тоническое сокращение гладкой мышечной ткани сосудов и полых внутренних органов, в том числе гладкомышечных сфинктеров. В этих органах содержатся и многочисленные рецепторные окончания в виде кустиков, деревцев или диффузных полей.
Регенерация гладкой мышечной ткани осуществляется несколькими способами:
· посредством внутриклеточной регенерациигипертрофии при усилении функциональной нагрузки;
· посредством митотического деления миоцитов при их повреждении (репаративная регенерация);
· посредством дифференцировки из камбиальных элементов - из адвентициальных клеток и миофибробластов.
Конец работы -
Эта тема принадлежит разделу:
ЛЕКЦИЯ 1. ВВЕДЕНИЕ В КУРС ГИСТОЛОГИИ 1. Определение гистологии как науки
ЛЕКЦИЯ ВВЕДЕНИЕ В КУРС ГИСТОЛОГИИ... Определение гистологии как науки Объекты исследования гистологии...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Все темы данного раздела:
Строение и функции плазмолеммы (цитолеммы)
Плазмолемма оболочка животной клетки, ограничивающая ее внутреннюю среду и обеспечивающая взаимодействие клетки с внеклеточной средой.
Плазмолемма имеет толщину около 10 нм, и состо
Строение и функции межклеточных контактов
В тех тканях, в которых клетки или их отростки плотно прилежат друг к другу (эпителиальная, гладкомышечная и другие) между плазмолеммами контактирующих клеток формируются связи - межклеточные конта
Гиалоплазма
Гиалоплазма или матрикс цитоплазмы составляет внутреннюю среду клетки. Она состоит из воды (90 %) и различных биополимеров (7 %) белков, нуклеиновых кислот, полисахаридов, липидов, из которых основ
Строение и функции общих органелл
Митохондрии наиболее обособленные структурные элементы цитоплазмы клетки, обладающие в значительной степени самостоятельной жизнедеятельностью. Существует даже точка зрения, что митохондрии
Строение и функции немембранных органелл
Рибосомы аппараты синтеза белка и полипептидных молекул. По локализации подразделяются на:
· свободныенаходятся гиалоплазме;
· несвободные или прикрепленныесвязаны с мембра
Реакция клеток на внешнюю среду
В организме человека содержатся только эукариотические (ядерные) типы клеток. Безъядерные структуры (эритроциты, тромбоциты, роговые чешуйки) являются вторичными (постклеточными) обра
Способы размножения (репродукции) клеток
Различают два основных способа размножения клеток:
· митоз (кариокенез) - непрямое деление клеток, которое присуще в основном соматическим клеткам;
· мейоз или редукционное делени
Реакция клеток на внешние воздействия
Описанная морфология клеток не является стабильной (постоянной). При воздействии на организм различных неблагоприятных факторов в строении различных структур проявляются различные изменения. В зави
Прогенез
Зрелые половые клетки, в отличие от соматических содержат одиночный (гаплоидный) набор хромосом. Все хромосомы гаметы, за исключением одной половой, называются аутосомами. В мужских п
Оплодотворение
Оплодотворение - процесс слияния мужской и женской гамет, приводящее к образованию зиготы. При оплодотворении взаимодействуют мужская и женская гаплоидные гаметы, при этом сливаются и
Формирования эмбриона и плодных оболочек
На 4-е сутки после оплодотворения, в полость матки выпадает морула. Морула - группа клеток, возникших в ходе нескольких делений дробления и заключенных внутри прозрачной оболочки. Около 2-х
Гистогенез и органогенез
Каждая клетка развивающегося зародыша содержит определенный набор геновгеном, совокупность генов организма - генотип.
В основе гистогенеза лежат следующие процессы:
Виды эпителиальных тканей
1. Ткань- исторически (филогенетически) сложившаяся система клеток и неклеточных структур, обладающая общностью строения, а иногда и происхождения, и специализированная на выполнен
Развитие тканей в онтогенезе (филогенезе)
В онтогенезе различают следующие этапы развития тканей:
· I этап топической дифференцировки - презумптивные (предположительные) зачатки тканей оказываются в определенных зонах цитоп
Регенерация тканей
Регенерация - восстановление клеток, направленное на поддержание функциональной активности данной системы. В регенерации различают такие понятия, как форма регенерации, уровень регенерации,
Интеграция тканей
Ткани, являясь одним из уровней организации живой материи, входят в состав структур более высокого уровня организации живой материи - структурно-функциональных единиц органов и в состав о
Функции и состав лимфы
1. Кровь и лимфа - это ткани внутренней среды организма, они является разновидностью соединительной ткани.
У данных видов тканей имеются следующие особенности: мезенхималь
Возрастные особенности крови
У новорожденных:
· эритроцитов 6-7 млн в 1 л (эритроцитоз);
· лейкоцитов 10-30 тыс. в 1 л (лейкоцитоз);
· тромбоцитов 200-300 тыс. в 1 л, то есть как у взрослых.
В-лимфоцитопоэз
1. Кроветворение(гемоцитопоэз)процесс образования форменных элементов крови.
Различают два вида кроветворения:
миелоидное кроветворение:
· эритроп
1. В понятие соединительные ткани (ткани внутренней среды, опорно-трофические ткани) объединяются неодинаковые по морфологии и выполняемым функциям ткани, но обладающие некоторыми
Структурная и функциональная характеристика клеточных типов
I. Фибробласты - преобладающая популяция клеток рыхлой волокнистой соединительной ткани. Они неоднородны по степени зрелости и функциональной специфичности и потому подразделяются на след
Межклеточное вещество соединительной ткани
Оно состоит из двух структурных компонентов:
· основного или аморфного вещества;
· волокон.
Основное или аморфное вещество состоит из белков и углеводов. Бе
Соединительные ткани со специальными свойствами
К ним относятся ретикулярная, жировая, слизистая и пигментная ткани.
Ретикулярная ткань состоит из ретикулярных клеток и ретикулярных волокон. Эта ткань образует строму всех кроветв
Остеогистогенез
1.К скелетным соединительным тканям относятся хрящевые и костные ткани, выполняющие опорную, защитную и механическую функции, а также принимающие участие в обмене минеральны
Развитие костной ткани и костей (остеогистогенез)
Все разновидности костной ткани развиваются из одного источника - из мезенхимы, но развитие разных костей осуществляется неодинаково. Различают два способа остеогистогенеза:
· разви
1.Свойством сократимости обладают практически все виды клеток, благодаря наличию в их цитоплазме сократительного аппарата, представленного сетью тонких микрофиламентов (5-7
Поперечно-полосатая скелетная мышечная ткань
Структурно-функциональной единицей поперечно полосатой мышечной ткани является мышечное волокно. Оно представляет собой вытянутое цилиндрическое образование с заостренными концами длиной от
Гистогенез и регенерация мышечной ткани
Из миотомов мезодермы в определенные участки мезенхимы выселяются малодифференцированные клетки - миобласты, часть из которых выстраивается в виде цепочки в стык друг к другу. В области конт
Иннервация и кровоснабжение скелетных мышц
Скелетные мышцы получают двигательную, чувствительную и трофическую (вегетативную) иннервацию. Двигательную (эфферентную) иннервацию скелетные мышцы туловища и конечностей получают от мотоне
Сердечная поперечно-полосатая мышечная ткань
Структурно-функциональной единицей является клетка -кардиомиоцит. По строению и функциям кардиомиоциты подразделяются на две основные группы:
· типичные или сократительные ка
Специальные гладкомышечные ткани
Нейрального происхождения развиваются из нейроэктодермы, из краев стенки глазного бокала, являющегося выпячиванием промежуточного мозга. Из этого источника развиваются миоциты, которые образ
Рецепторные нервные окончания
1.Значение нервной ткани в организме определяется основными свойствами нервных клеток (нейронов, нейроцитов) воспринимать раздражение, приходить в состояние возбуж
Нейроглия
Классификация нейроглии:
· макроглия (глиоциты):
· эпендимоциты;
· астроциты;
· олигодендроциты;
· микроглия.
Эпендимоциты образу
Нервные волокна
Отростки нервных клеток, обычно покрытые оболочками, называются нервными волокнами. В различных отделах нервной системы оболочки нервных волокон значительно отличаются друг от друга по своему строе
Регенерация нейронов и нервных волокон
Нейроны являются несменяемой клеточной популяцией. Им свойственна только внутриклеточная физиологическая регенерация, заключающаяся в непрерывной смене структурных белков цитоплазмы.
Отрос
Рецепторные нервные окончания
Главная функция афферентных нервных окончаний является восприятие сигналов поступающих из внешней и внутренней среды. Рецептор - это терминальное ветвление дендрита чувствительной (рецепторн
Мозговые оболочки
1. Нервная система осуществляет объединение частей организма в единое целое (интеграцию), обеспечивает регуляцию разнообразных процессов, координацию функции различных орган
Ствол головного мозга
Головной мозг состоит из ствола мозга, который является продолжением спинного мозга (включает продолговатый, задний, средний и промежуточный мозг) и плащевой части, образованной полушариями
Миелоархитектоника и организация коры
Нервные волокна коры полушарий большого мозга включают три группы:
· афферентные;
· ассоциативные и комиссуральные;
· эфферентные волокна.
Афферентные
Мозговые оболочки
Головной мозг защищен костями черепа, а спинной - позвонками и межпозвонковыми дисками; они окружены тремя мозговыми оболочками (снаружи внутрь): твердой, паутинной и мягкой, которые фиксируют эти
Производные кожи
1. Кожа покрывает поверхность тела и является одним из наиболее крупных органовее масса составляет около 16 % массы тела, а площадь поверхности 1,2 2,3 кв. м. К производным кожи у
Железы кожи
Потовые железы участвуют в терморегуляции, а также в экскреции продуктов обмена, солей, лекарственных веществ, тяжелых металлов (усиливается при почечной недостаточности). Потовые железы под
Производные кожи
Волосы - ороговевшие нитевидные придатки кожи толщиной 0,005-0,6 мм и длинной от нескольких миллиметров до 1,5 м, их цвет, размеры и распределение связаны с возрастом, полом, расовой принадл
Аккомодационный аппарат глаза
1. Сенсорная система обеспечивает восприятие организмом информации о состоянии внешней и внутренней среды, а также ее обработку и трансформацию в ощущения. Все эти функции о
Гистофизиология слуха
Звуки определенной частоты воспринимаются наружным ухом и передаются через слуховые косточки и овальное окно перилимфе в барабанной и вестибулярной лестницах. При этом приходят в колебательн
Диоптрический аппарат глаза
Роговица - прозрачная часть наружной фиброзной оболочки глаза склеры. Она состоит из пяти слоев:
· наружный эпителий является многослойным плоским неороговевающим эпителием,
Аккомодационный аппарат глаза
Сосудистая оболочка состоит из трех частей: собственно сосудистой оболочки, цилиарного тела и радужки. Главная функция собственно сосудистой оболочки - питание сетчатки. Она также участвует
Лимфатические сосуды
1. Сердечно-сосудистая система образована сердцем, кровеносными и лимфатическими сосудами.
Функции сердечно-сосудистой системы:
· транспортная - обеспечени
Кровоснабжение легких
1. Дыхательная система состоит из двух частей: воздухоносных путей и респираторного отдела. К воздухоносным путям относят полость носа, носоглотку, трахею, бронхиа
Полость носа
Полость носа состоит из преддверия и дыхательной части. Преддверие носа выстлано слизистой оболочкой, в составе которой находится многослойный плоский неороговевающий эпителий и собственная
Строение паращитовидной железы
1. Эндокринная система относится к числу регуляторно-интегрирующих систем организма наряду с сердечно-сосудистой, нервной и иммунной, выступая с ними в теснейшем единстве. В ее вед
Гипофиз
Аденогипофиз развивается из эпителия крыши ротовой полости, имеющей эктодермальное происхождение. На 4-й неделе эмбриогенеза образуется эпителиальное выпячивание этой крыши в виде кармана Ра
Паращитовидные железы
Основная функция паращитовидных желез - секреция гормонов:
· гормон паратирин, который является антагонистом тирокальцитонина, он повышает уровень кальция в крови двумя способами
Строение поджелудочной железы
1. Пищеварительная система обеспечивает поступление в организм питательных веществ и расщепление их до мономеров, способных всасываться в кровь и лимфу, а также выведение нерасщепл
Развитие зуба
Основными источниками развития зубов являются эпителий слизистой оболочки ротовой полости (эктодерма) и мезенхима. У человека различают две генерации зубов: молочные и постоянные. Их развити
Железы языка
Язык содержит большое количество слюнных желез. Их концевые отделы лежат в прослойках рыхлой волокнистой соединительной ткани между мышечными волокнами и в подслизистой оболочке нижней поверхности.
Желудок
К среднему отделу пищеварительного тракта относятся желудок и кишечник за исключением анального отдела прямой кишки. В состав этих органов входит слизистая оболочка кишечного типа, выстланная однос
Функции аппендикса
1. Все органы кроветворения и иммуногенеза делятся на:
· центральные - красный костный мозг, тимусдля Т-лимфопоэза;
· периферические - лимфоузлы,
Миндалины
В отличии от лимфоузлов и селезенки, относящихся к так называемым лимфоретикулярным органам иммунной системы, миндалины называют лимфоэпителиальными органами. Так как в них осуществля
Мочевыводящие пути
Мочевыделительная система состоит из почек - мочеобразующего органа и мочевыводящих путей: почечных лоханок и чашечек, мочеточников, мочевого пузыря и мочеиспускательного канала.
Гистофизиология нефрона
Структурно-функциональной единицей почки является нефрон. Он состоит из капсулы и переходящих друг в друга канальцевпроксимальных извитого и прямого, дистальных извитого и прямого. В каждой
Кровоснабжение почки
Сосуды почки имеют характерную архитектонику в связи с наличием двух основных видов нефронов: корковых и юкстамедуллярных.
Кровь поступает в почку через почечную артерию, которая де
Добавочные железы
1. Мужская половая система выполняет важные функции: обеспечивает полноценное развитие половых клеток, их кондиционирование (окончательное структурное и функциональное созревание)
Функции семявыносящих путей
Семявыносящие пути выполняют следующие функции:
· депонирование, трофика, кондиционирование спермы;
· обеспечение массированного выброса спермы во время коитуса;
·
Добавочные органы и железы
К этим органам относятся семенные пузырьки, предстательная железа, бульбоуретральные железы, половой член. Функциями этих образований являются участие в кондиционировании спермы, трофике сперматозо
Менструальный цикл
1.В отличие от мужской женская половая система обеспечивает не только образование половых клеток и синтез гормонов, но и вынашивание и вскармливание потомства. В с
Строение яйцеводов и матки
Анатомически яйцеводы состоят из воронковой, ампулярной, истмической и маточной частей. Они являются органами слоистого типа, стенка образована тремя оболочками: слизистой, мышечной и серозн
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
РАЗВИТИЕ
Источником развития скелетной мышечной ткани являются миотомы сомитов . Стадии развития:
- Миобластическая стадия . Клетки миотомов превращаются в миобласты и мигрируют к местам закладки мышц. Миобласты делятся митозом. Часть миобластов обособляется в виде миосателлитоцитов, которые сохраняют свойства малодифференцированных клеток до конца жизни.
- Миосимпластическая стадия . Миобласты располагаются в виде цепочек и сливаются друг с другом. Образуются миосимпласты. В их цитоплазме образуются миофибриллы из сократительных белков, которые начинают синтезироваться ещё в миобластах. Миофибриллы лежат на периферии миосимласта, ядра занимают центральное положение.
- Стадия миотубул . В симпластах увеличивается число миофибрилл. Длина их увеличивается.
- Стадия зрелого мышечного волокна . В эту стадию объём миофибрилл увеличивается до такой степени, что они занимают основную массу волокна, смещаясь в центр и сдвигая ядра на периферию.
СТРОЕНИЕ МЫШЕЧНОГО ВОЛОКНА .
Мышечные волокна являются структурно-функциональным элементом скелетной мышечной ткани. Они имеют длину до 20-30 см, толщину около 100 мкм. Состоят из двух частей:
· симпласта;
· миосателлитоцитов.
Мышечное волокно снаружи покрыто сарколеммой . Сарколемма состоит из толстой базальной мембраны и плазмолеммы мышечного волокна. Между базальной мембраной и плазмолеммой в отдельных участках имеются углубления, в которых расположены миосателлитоциты . Миосателлитоциты – камбиальные клетки скелетной мышечной ткани. Миосателлитоцит – типичная одноядерная клетка, окруженная своей плазмолеммой, имеет слабо развитые органеллы.
Миосимпласт содержит множество (до несколько тысяч) ядер, лежащих на периферии волокна, под сарколеммой.
Протоплазму волокна называют саркоплазмой. В ней находятся органеллы общего значения (за исключением центриолей), органеллы специального значения – миофибриллы , и включения.
СТРОЕНИЕ МИОФИБРИЛЛ .
Миофибриллы , числом до двух тысяч в одном волокне, занимают основную часть волокна. Их длина равна длине волокна, диаметр до 2 мкм. В каждой миофибрилле при световой микроскопии обнаруживается исчерченность – чередование светлых и тёмных дисков. В поляризованном свете тёмные диски имеют двойное лучепреломление и поэтому тёмные диски называются анизотропными , или А-дисками . Светлые диски не имеют двойного лучепреломления и называются изотропными , или I-дисками .
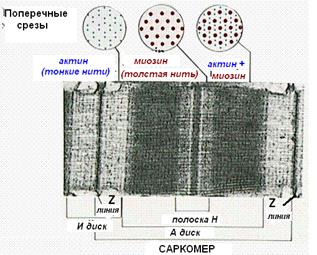
Посередине I-диска проходит тёмная полоска, которая называется Z-линией, или телофрагмой. На поперечном разрезе телофрагма представляет собой решётку, в узлах которой закрепляются актиновые филаменты.
В центре А-диска находится более светлая полоска Н, а посередине её проходит тёмная линия М , или мезофрагма .
Участок миофибриллы, лежащий между соседними Z-линиями, называется саркомером. Саркомер – структурно-функциональная единица миофибриллы. В состав саркомера последовательно входят:
· Z-линия;
· ½ диска I;
· диск А;
· ½ диска I
· вторая Z-линия.
Каждый саркомер состоит из тонких актиновых и толстых миозиновых филаментов.
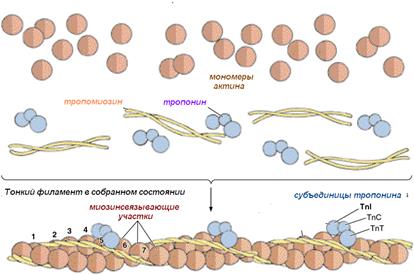
В составе тонких (диаметр 5 нм) актиновых филаментов входят белки:
· актин;
· тропонин;
· тропомиозин .
Молекулы актина имеют глобулярное строение – G-актин. Эти молекулы соединяются вместе в длинные цепочки – фибриллярный, F-актин . В актиновых филаментах две цепи F-актина образуют двойную спираль. В бороздках между цепями спирали лежат молекулы тропомиозина . К молекулам тропомиозина на равных расстояниях друг от друга прикрепляются молекулы тропонина. Молекула тропонина состоит из трёх субъединиц: TnT, TnI, TnC . TnT осуществляет прикреплениетропонина к тропомиозину.TnC отвечает за связывание с ионами кальция. TnI препятствует взаимодействию миозина с актином.
Толстые филаменты (диаметр 12 нм) содержат белок миозин . Каждая молекула миозина состоит из двух частей: головки и хвоста и может сгибаться в двух местах – шарнирных участках. Головка миозина имеют АТФ-азную активность и способна расщеплять АТФ с образованием энергии. Молекулы миозина соединяются в пучки и образуют толстые миозиновые филаменты. По периферии толстых филаментов находятся участки, содержащие головки миозина. Центральная часть не содержит головок.
В составе саркомера толстые филаменты лежат только в диске А. Тонкие филаменты расположены в дискеI, но концами частично заходят в диск А между миозиновыми филаментами. Та часть диска А, которая содержит и актиновые и миозиновые филаменты, выглядит на срезах более тёмной, а та его часть, которая содержит только миозиновые филаменты, светлее. Эта часть диска А, которая содержит только миозиновые филаменты, и составляет полоску Н. Таким образом:
· диск I состоит из актиновых филаментов;
· полоска Н диска А состоит из миозиновых филаментов;
· на периферии диска А есть зона пересечения актиновых и миозиновых филаментов.
На поперечном срезе миофибриллы можно видеть, что в зоне пересечения вокруг одной толстой филаменты лежат шесть тонких филамент.
Тонкие филаменты неподвижно прикреплены к Z-линиям. В состав Z-линий входят белки α-актинин, десмин, виментин.
Линия М в центре Н-полоски – место соединения всех миозиновых филаментов друг с другом. В их скреплении участвуют белки миомезин и С-белок.
Физиология скелетных мышц
Физическая нагрузка реализуется в результате согласованных действий скелетной мускулатуры. Рассмотрим основные характеристики их структуры и функции.
Взаимодействие человека с внешней средой не может осуществляться без сокращений его мышц. Производимые при этом движения необходимы как для выполнения простейших манипуляций, так и для выражения самых тонких мыслей и чувств - посредством речи, письма, с помощью мимики или жестов. Масса мышц намного больше, чем других органов; они составляют 40-50% массы тела. Мышцы - это «машины», преобразующие химическую энергию непосредственно в механическую (работу) и в теплоту. Деятельность их, в частности механизм укорочения и генерирования силы, сейчас можно достаточно детально объяснить на молекулярном уровне с использованием физических и химических законов.
Рис 1. Структура скелетных мышц: организация цилиндрических волокон в скелетной мышце, прикрепленной к костям сухожилиями.
Понятие скелетная, или поперечно-полосатая мышца относится к группе мышечных волокон, связанных соединительной тканью (рис. 1 ). Обычно мышцы прикреплены к костям пучками коллагеновых волокон - сухожилиями, находящимися на обоих концах мышцы. В некоторых мышцах одиночные волокна имеют такую же протяженность, как и вся мышца, но в большинстве случаев волокна короче и часто располагаются под углом к продольной оси мышцы. Есть очень длинные сухожилия, они прикреплены к кости, удаленной от конца мышцы. Например, некоторые мышцы, осуществляющие движения пальцев рук, находятся в предплечье; шевеля пальцами, мы чувствуем, как двигаются мышцы кисти. Эти мышцы соединены с пальцами посредством длинных сухожилий
Что такое скелетная мышца?
Один грамм ткани скелетной мышцы содержит примерно 100 мг «сократительных белков» - актина (молекулярная масса 42000) и миозина (молекулярная масса 500 000).
Скелетная мышца, например, бицепс, по виду кажется единым образованием, но на самом деле состоит из тканей нескольких типов. В состав каждой мышцы входят длинные тонкие цилиндрические мышечные волокна (клетки), вытянутые по всей ее длине; поэтому они могут быть очень длинными. Каждая многоядерная мышечная клетка (волокно) окружена параллельными мышечными волокнами, с которыми связана слоем соединительной ткани под названием эндомизий. Эти волокна собраны в пучки, скрепленные слоем соединительной ткани, под названием перимизий. Такая упакованная группа, или связка, волокон называется мышечным пучком. Группы пучков с прилегающими к ним сосудами и нервами связаны друг с другом с помощью еще одного слоя соединительной ткани под названием эпимизий. Собранные вместе и окруженные эпимизием пучки, которые тянутся по всей длине скелетной мышцы, покрыты сверху слоем соединительной ткани, называемым фасцией.
Какова функция фасции в скелетной мышце?
Фасция - это упругая, плотная и прочная соединительнотканная оболочка, которая покрывает мышцу целиком и, выходя за ее пределы, образует фиброзное сухожилие. Фасция образована посредством слияния всех трех внутренних слоев соединительной ткани скелетной мышцы. Фасция отделяет мышцы друг от друга, уменьшает трение при движении и формирует сухожилие, с помощью которого мышца прикрепляется к костному скелету. Этому компоненту мышц обычно не уделяется должного внимания. Тем не менее, многие специалисты полагают, что для свободного неограниченного движения мышцы, а, следовательно, и сустава совершенно необходимо свободное движение фасции.
Рис. 2. Структура скелетных мышц: структурная организация филаментов в волокне скелетной мышцы, создающая картину поперечных полос.
Почему скелетная мышца называется поперечно-полосатой?
При изучении с помощью светового микроскопа, основной характеристикой волокон скелетной мышцы оказалось чередование светлых и темных полос, поперечных по отношению к длинной оси волокна. Поэтому скелетные мышцы были названы поперечно-полосатыми.
Поперечная исчерченность волокон скелетной мышцы обусловлена особым распределением в их цитоплазме многочисленных толстых и тонких «нитей» (филаментов), объединяющихся в цилиндрические пучки диаметром 1-2 мкм - миофибриллы (рис. 2 ). Мышечное волокно практически заполнено миофибриллами, они тянутся по всей его длине и на обоих его концах соединены с сухожилиями. Миофибриллы состоят из сократительных филаментов (белков). Основных сократительных микро-филаментов два - миозин и актин. Структурное расположение этих белков придает скелетной мышце вид перемежающихся светлых и темных полос. Каждая темная полоса (полоса, или диск, А) соответствует участку, где актиновые и миозиновые белки перекрываются, тогда как более светлая полоса - участку, где их перекрытия нет (полоса, или диск, I). Перегородки, называемые Z-пластинками, разделяют их на несколько компартментов-саркомеров - длиной примерно по 2,5 мкм.
Что является структурной единицей мышечной ткани скелетной мышцы?
Структурной единицей мышечной ткани скелетной мышцы являются мышечные клетки, которые существенно отличаются от других мышечных тканей, прежде всего от гладких мышц
Гладкое мышечное волокно - это веретеновидная клетка диаметром от 2 до 10 мкм. В отличие от многоядерных волокон скелетных мышц, которые после завершения дифференцировки уже не могут делиться, гладкие мышечные волокна обладают единственным ядром и способны к делению на протяжении всей жизни организма. Деление начинается в ответ на разнообразные паракринные сигналы, часто - на повреждение ткани.
Поперечно-полосатые мышцы скелета состоят из множества функциональных единиц - мышечных волокон, которые расположены в общем соединительнотканном футляре. Каждое волокно скелетной мышцы - это тонкое (диаметром 0,01-0,1 мм), вытянутое на 2-3 см, многоядерное образование - симпласт-результат слияния многих клеток. Ядра в волокне расположены около его поверхности. Пучки мышечных волокон окружены коллагеновыми волокнами и соединительной тканью; между волокнами также находится коллаген. На конце мышц коллаген вместе с соединительной тканью образует сухожилия, которые служат для прикрепления мышц к разным частям скелета. Каждое волокно окружено мембраной - сарколеммой, которая по своему строению сходна с плазматической мембраной.
Основной особенностью мышечного волокна является наличие в его цитоплазме - саркоплазме большого количества тонких нитей - миофибрилл, расположенных вдоль оси волокна. Миофибриллы состоят из чередующихся светлых и темных участков - дисков, что придает мышечному волокну поперечную исчерченность (полосатость).
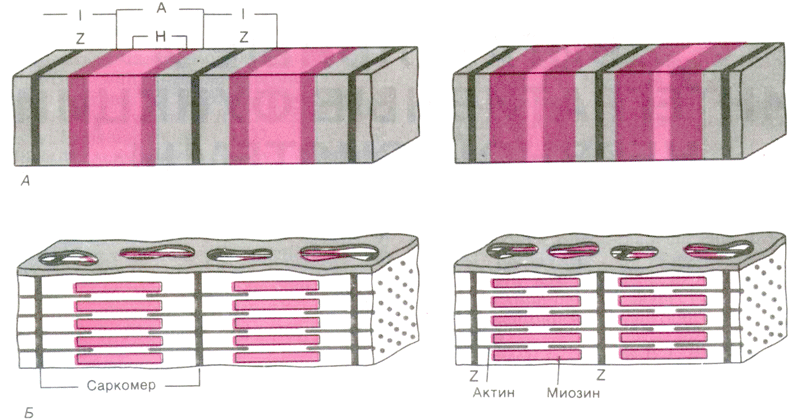
Рис 3. Организация миозиновых и актиновых нитей в расслабленном и сократившемся саркомере.
Что такое саркомер?
Это минимальная сократительная единица скелетной мышцы.
Рассмотрим более детальноструктуру саркомеров, которая схематически показана на рис 3 . С помощью светового микроскопа в них можно видеть регулярно чередующиеся поперечные светлые и темные полосы. Согласно теории Хаксли и Хансон, такая поперечная полосатость миофибрилл обусловлена особым взаиморасположением актиновых и миозиновых филаментов. Середину каждого саркомера занимают несколько тысяч «толстых» нитей миозина диаметром примерно по 10 нм. На обоих концах саркомера находятся около 2000 «тонких» (толщиной по 5 нм) нитей актина, прикрепленных к Z-пластинкам наподобие щетинок в щетке.
Толстые филаменты сосредоточены в средней части каждого саркомера, где они лежат параллельно друг другу; эта область выглядит как широкая темная (анизотропная) полоса, называемая А-полосой. В обеих половинах саркомера находится по набору тонких филаментов. Один конец каждого из них прикреплен к так называемой Z-пластинке (или Z-линии, или Z-полосе) - сети из переплетающихся белковых молекул, - а другой конец перекрывается с толстыми филаментами. Саркомер ограничен двумя последовательно расположенными Z-полосами. Таким образом, тонкие филаменты двух соседних саркомеров закреплены на двух сторонах каждой Z-полосы.
В пределах А-полосы каждого саркомера различают еще две полоски. В центре А-полосы видна узкая светлая полоска - Н-зона. Она соответствует промежутку между противостоящими друг другу концами двух наборов тонких филаментов каждого саркомера, т.е. включает только центральные части толстых филаментов. Посередине Н-зоны находится совсем тонкая темная М-линия. Это сеть белков, соединяющих центральные части толстых филаментов. Кроме того, от Z-полосы к М-линии идут филаменты белка титина, связанные одновременно с белками М-линии и с толстыми филаментами. М-линия и титиновые филаменты поддерживают упорядоченную организацию толстых филаментов в середине каждого саркомера. Таким образом, толстые и тонкие филаменты не являются свободными, незакрепленными внутриклеточными структурами.
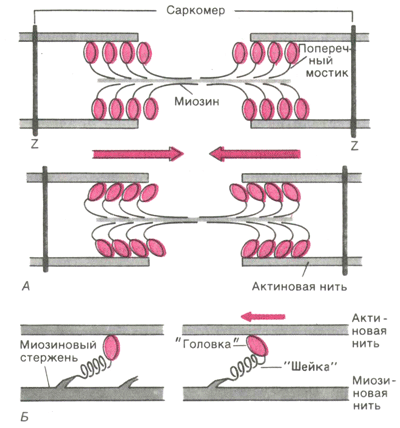
Рис 4. Функция поперечных мостиков. А. Модель механизма сокращения
Обсудим собственно механизм мышечного сокращения
Каким образом взаимодействуют актин и миозин?
Активные участки молекулы актина, способные связывать глобулярные головки миозина, располагаются на ней на некотором расстоянии друг от друга. Когда эти активные участки открыты, миозиновая головка самопроизвольно связывается с актиновым филаментом и образует поперечный мостик. При снабжении миозиновой головки достаточным количеством энергии глобулярная головка подтягивает актин по направлению к центру саркомера, что часто называют храповым движением. Это движение укорачивает саркомер.
Работа поперечных мостиков (Рис. 4) . Во время сокращения каждая миозиновая головка может связывать миозиновую нить с соседними актиновыми. Движение головок создает объединенное усилие, как бы «гребок», продвигающий актиновые нити к середине саркомера. Сама биполярная организация молекул миозина обеспечивает противоположную направленность скольжения актиновых нитей в левой и правой половинах саркомера. В результате однократного движения поперечных мостиков вдоль актиновой нити саркомер укорачивается только на 2 х 10 нм, т. е. примерно на 1% своей длины. За счет ритмичных отделений и повторных прикреплений миозиновых головок актиновая нить может подтягиваться к середине саркомера, подобно тому, как группа людей тянет длинную веревку, перебирая ее руками. Следовательно, когда принцип «вытягивания веревки» реализуется во множестве последовательных саркомеров, повторяющиеся молекулярные движения поперечных мостиков приводят к макроскопическому движению. При расслаблении мышцы миозиновые головки отделяются от актиновых нитей. Поскольку актиновые и миозиновые нити могут легко скользить друг относительно друга, сопротивление расслабленных мышц растяжению очень низкое. Их можно снова растянуть до исходной длины, приложив совсем небольшое усилие. Следовательно, удлинение мышцы во время расслабления носит пассивный характер.
Рис 5. Функция поперечных мостиков. Б. Модель механизма генерирования силы поперечными мостиками: слева до, справа - после «гребка
Генерирование мышечной силы . Благодаря упругости поперечных мостиков саркомер может развивать силу даже без скольжения нитей относительно друг друга, т. е. в строго изометрических экспериментальных условиях. Рис.5.Б иллюстрирует такой процесс генерирования изометрической силы. Сначала головка миозиновой молекулы прикрепляется к актиновой нити под прямым углом. Затем она наклоняется под углом примерно 45°, возможно, благодаря притяжению между соседними точками прикрепления на ней и на актиновой нити. При этом головка действует как миниатюрный рычаг, приводя внутреннюю упругую структуру поперечного мостика (видимо, «шейки» между головкой и миозиновой нитью) в напряженное состояние. Возникающее в результате упругое растяжение достигает лишь около 10 нм. Упругое натяжение, создаваемое индивидуальным поперечным мостиком, так слабо, что для развития мышечной силы, равной 1 мН, нужно объединить усилия, по крайней мере, миллиарда таких соединенных параллельно мостиков. Они будут тянуть соседние актиновые нити, как команда игроков тянет канат. Даже при изометрическом сокращении поперечные мостики не находятся в непрерывно напряженном состоянии (это наблюдается только при трупном окоченении). На самом деле каждая миозиновая головка уже через сотые или десятые доли секунды отделяется от актиновой нити; однако через такое же короткое время следует новое прикрепление к ней. Несмотря на ритмичное чередование прикреплений и отделений с частотой порядка 5 - 50 Гц, сила, развиваемая мышцей в физиологических условиях, остается неизменной (исключение - летательные мышцы насекомых), так как статистически в каждый момент времени в прикрепленном, обусловливающем напряжение, состоянии находится одно и то же количество мостиков.
Что такое цикл поперечного мостика?
Цикл поперечного мостика - это термин, описывающий взаимодействие глобулярной головки миозина с активным участком молекулы актина. Формированию поперечного мостика способствуют два фактора: повышение внутриклеточной концентрации ионов кальция и присутствие аденозинтрифосфата (АТФ). Один цикл поперечного мостика состоит из:
активации миозиновой головки;
обнажения активного участка молекулы актина в присутствии кальция;
самопроизвольного формирования поперечного мостика;
поворота глобулярной головки, сопровождающегося продвижением актиновой нити и укорочением саркомера;
отцепления поперечного мостика.
Цикл после завершения может повторяться или останавливаться. Поворот миозиновой головки называется также рабочим ходом.
Чем предотвращается спонтанное взаимодействие миозина и актина после отцепления поперечного мостика? Каков механизм циклического формированию поперечного мостика – повторяющегося взаимодействия глобулярной головки миозина с активным участком молекулы актина?
Для понимания всего этого необходимо внимательнее рассмотреть строение миозина и, особенно, актина.
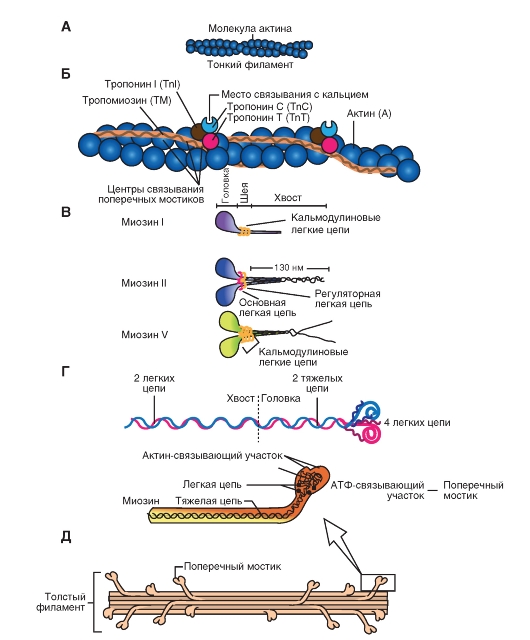
Рис. 6. Строение миозина
Это единое название большой семьи протеинов, имеющих определенные отличия в клетках разных тканей. Миозин присутствует у всех эукариотов. Около 60 лет назад было известно два типа миозина, которые сейчас называют миозин I и миозин II. Миозин II был первым из числа открытых миозинов, и именно он принимает участие в мышечном сокращении. Позднее были открыты миозин I и миозин V (рис. 6 В ). В последнее время показано, что миозин II участвует в мышечном сокращении, тогда как миозин I и миозин V вовлечены в работу подмембранного (кортикального) цитоскелета. В настоящее время идентифицировано более 10 классов миозина. На рисунке 6 Г показано два варианта схемы строения миозина, который состоит из головки, шейки и хвоста. Молекула миозина состоит из двух больших полипептидов (тяжелых цепей) и четырех меньших (легких цепей). Эти полипептиды составляют молекулу с двумя глобулярными «головками», которые содержат оба вида цепей, и длинным стержнем («хвостом») из двух переплетенных тяжелых цепей. Хвост каждой молекулы миозина располагается вдоль оси толстого филамента, а две глобулярные головки выступают по бокам.На каждой глобулярной головке находятся по два участка связывания: для актина и для АТФ. Участки связывания АТФ обладают также свойствами фермента АТФазы, гидролизующей связанную молекулу АТФ.
Рис 7. Строение актина
Молекула актина
Это глобулярный белок, состоящий из одного полипептида, который полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга (рис. 7 А ). Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся мышечном волокне взаимодействие между актином и миозином предотвращают два белка - тропонин и тропомиозин (рис. 7 Б ).
Тропонин - гетеротримерный белок. Он состоит из тропонина Т (отвечает за связывание с одиночной молекулой тропомиозина), тропонина С (связывает ион Са 2+) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
Чем предотвращается спонтанное взаимодействие миозина и актина?
В желобках двойной спирали актина располагаются два дополнительных регуляторных белка, которые предотвращают самопроизвольное взаимодействие актина и миозина. Эти белки, тропонин и тропомиозин, играют важную роль в процессе сокращения скелетной мышцы. Функция тропомиозина заключается в том, что в покое он закрывает (защищает) активные участки актинового филамента. Тропонин имеет три связывающих участка: один служит для связывания ионов кальция (тропонин С), другой прочно прикреплен к молекуле тропомиозина (тропонин Т), третий связан с актином (тропонин I). В покое эти регуляторные белки закрывают связывающие участки на молекуле актина и препятствуют формированию поперечных мостиков. Все эти микроструктурные компоненты вместе с митохондриями и другими органеллами клетки окружены клеточной мембраной, называемой сарколеммой.
![]()
Рис. 8. Действие Са 2+ во время активации миофибриллы.
А.Актиновая и миозиновая нити на продольном сечении волокна. Б. Они же на его поперечном сечении.
Исследования с помощью рентгеноструктурного анализа (малоугловое рентгеновское рассеяние) показали, что в отсутствие Са 2+ , т. е. при расслабленном состоянии миофибрилл, длинные молекулы тропомиозина располагаются так, что блокируют прикрепление поперечных миозиновых головок к актиновым нитям. И напротив, когда Са 2+ связывается с тропонином, тропомиозин попадает в желобок между двумя мономерами актина, обнажая участки прикрепления для поперечных мостиков (Рис . 8).
Если активные участки закрыты, то каким образом взаимодействуют актин и миозин?
Когда внутри клетки повышается концентрация ионов кальция, они связываются с тропонином С. Это приводит к изменениям конформации тропонина. В результате изменяется также трехмерная структура тропомиозина и обнажается активный участок молекулы актина. Сразу после этого головка миозина самопроизвольно связывается с активным участком актинового филамента, образуя поперечный мостик, который начинает двигаться и способствует укорочению саркомера. Наличие или отсутствие в клетке кальция частично регулируется сарколеммой (специализированной клеточной мембраной скелетной мышцы).
Какова функция кальция в скелетных мышцах?
Кальций обеспечивает открытие участков актиновой нити, связывающих миозин. Ионы кальция внутри клетки хранятся в СР(саркоплазматическом ретикуломе) и высобождаются после деполяризующей стимуляции. После высвобождения кальций диффундирует и связывается с белком - тропонином С. В результате конформация белка изменяется, он тянет молекулу тропомиозина и обнажает активные участки молекулы актина. Активные участки остаются открытыми все время, пока продолжается связывание кальция с тропонином С.
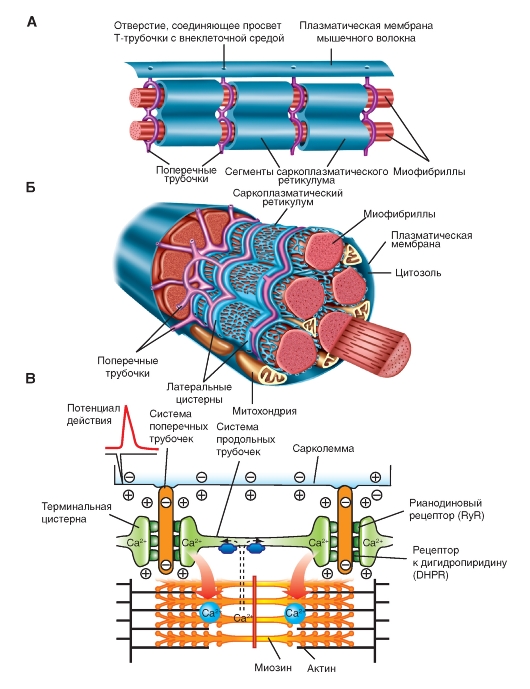
Рис. 9. Схема организации саркоплазматического ретикулума, поперечных трубочек и миофибрилл.
Хранение и высвобождение ионов кальция . Расслабленная мышца содержит более 1 мкмоль Са 2+ на 1 г сырой массы. Если бы соли кальция не были изолированы в особых внутриклеточных хранилищах, обогащенные его ионами мышечные волокна находились бы в состоянии непрерывного сокращения.
Источником поступления Са 2+ в цитоплазму служит саркоплазматический ретикулум мышечного волокна.
Саркоплазматический ретикулум мышц гомологичен эндоплазматическому ретикулуму других клеток. Он располагается вокруг каждой миофибриллы наподобие «рваного рукава», сегментами которого окружены А- и I-полосы (Рис. 9). Концевые части каждого сегмента расширяются в виде так называемых латеральных мешков (терминальных цистерн), соединенных друг с другом серией более тонких трубок. В латеральных мешках депонируется Са 2+ , высвобождающийся после возбуждения плазматической мембраны (рис. 10 ).
Рис. 10. Схема анатомической структуры поперечных трубочек и саркоплазматического ретикулума в индивидуальном волокне скелетной мышцы
Что такое поперечные трубочки (Т-трубочки) ?
Инвагинации на поверхности сарколеммы, расположенные на некотором расстоянии друг от друга. Благодаря Т-трубочкам внеклеточная жидкость может тесно контактировать с внутренними микроструктурами клетки. Т-трубочки представляют собой продолжение сарколеммы и также способны передавать потенциал действия на внутреннюю поверхность клетки. С Т-трубочками тесно взаимодействует саркоплазматический ретикулум (СР).
Что такое саркоплазматический ретикулум?
Специализированный эндоплазматический ретикулум, который состоит из везикул, ориентированных вдоль сократительных волокон скелетной мышцы. Эти везикулы осуществляют хранение, высвобождение во внутриклеточную жидкость и обратный захват ионов кальция. Специализированные расширенные участки СР называются концевыми цистернами. Концевые цистерны находятся в непосредственной близости от Т-трубочки и вместе с СР составляют структуру под названием триада. Особенности строения сарколеммы и триад играют важную роль в обеспечении саркомера ионами кальция, необходимыми для цикла поперечных мостиков.
Рис. 11. Роль саркоплазматического ретикулума в механизме сокращения скелетной мышцы
Возникнув в плазматической мембране (рис. 11 ), потенциал действия быстро распространяется по поверхности волокна и по мембране Т-трубочек вглубь клетки. Достигнув области Т-трубочек, прилегающей к латеральным мешкам, потенциал действия активирует потенциалзависимые «воротные» белки мембраны Т-трубочек, физически или химически сопряженные с кальциевыми каналами мембраны латеральных мешков. Таким образом, деполяризация мембраны Т-трубочек, обусловленная потенциалом действия, приводит к открыванию кальциевых каналов мембраны латеральных мешков, содержащих Са 2+ в высокой концентрации, и ионы Са 2+ выходят в цитоплазму. Повышение цитоплазматического уровня Са 2+ обычно бывает достаточным для активации всех поперечных мостиков мышечного волокна.
Процесс сокращения продолжается, пока ионы Са 2+ связаны с тропонином, т.е. до тех пор, пока их концентрация в цитоплазме не вернется к низкому исходному значению. Мембрана саркоплазматического ретикулума содержит Са-АТФазу - интегральный белок, осуществляющий активный транспорт Са 2+ из цитоплазмы обратно в полость саркоплазматического ретикулума. Как только что говорилось, Са 2+ высвобождается из ретикулума в результате распространения потенциала действия по Т-трубочкам; для возвращения Са 2+ в ретикулум нужно гораздо больше времени, чем для его выхода. Именно поэтому, повышенная концентрация Са 2+ в цитоплазме сохраняется в течение некоторого времени, и сокращение мышечного волокна продолжается после завершения потенциала действия.
Подведем итог. Сокращение обусловлено высвобождением ионов Са 2+ , хранящихся в саркоплазматическом ретикулуме. Когда Са 2+ поступает обратно в ретикулум, сокращение заканчивается и начинается расслабление.
Каковы особенности сарколеммы?
Электрический заряд на сарколемме, как и на других селективно проницаемых и возбудимых мембранах, образуется вследствие неодинакового распределения ионов. Проницаемость сарколеммы изменяется при стимуляции ацетилхолиновых рецепторов, расположенных в нервно-мышечном соединении. После достаточной стимуляции сарколемма может проводить деполяризующий сигнал (потенциал действия) по всей своей длине, а также в уникальную проводящую систему Т-трубочек.
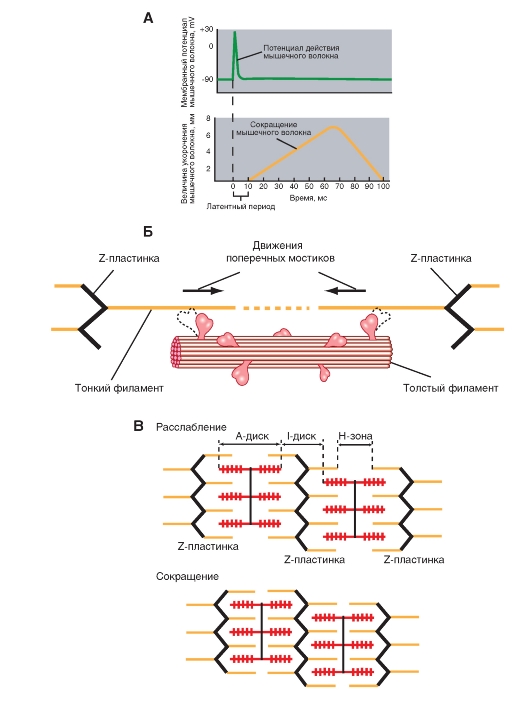
Рис. 12. Феномен электромеханического сопряжения


