Половое и бесполое размножение живых организмов. Научная библиотека - рефераты - способы размножения живых организмов. эволюция размножения
Размножение – это способность организмов производить себе подобных представителей того же вида, обеспечивая непрерывность и преемственность жизни в ряду поколений.. В процессе размножения особи родительского поколения передают потомкам генетическую информацию, обеспечивающую воспроизведение у них как признаков конкретных родителей, так и вида, к которому они принадлежат. Благодаря размножению осуществляется смена и материальная преемственность поколений. В ходе размножения создаются уникальные комбинации наследственного и закрепляются наследственные изменения, возникающие у отдельных особей. Это, в свою очередь, обуславливает генетическое разнообразие особей в пределах вида и служит основой для изменчивости и дальнейшей эволюции вида. Таким образом, осуществляемая в ходе размножения смена поколений, служит непременным условием поддержания во времени биологических видов и жизни.
Известны две формы размножения: бесполое и половое. При бесполом размножении особь появляется из неспециализированных (соматических) клеток тела. При половом размножении смена поколений и развитие организмов происходит на основе слияния специализированных клеток (половых) и образования зиготы. В природе существуют разные варианты двух основных форм размножения.
Типы размножения
Бесполое Половое
Одноклеточные Многоклеточные С оплодотворением Нерегулярные
Митоз Спорообразование Изогамия Партеногенез
Спорообразование Почкование Гетерогамия Гиногенез
Почкование Вегетативное Оогамия Андрогенез
Шизогония Фрагментация
Полиэмбриония
шизогония
2. Бесполое размножение.
Осуществляется при участии одной родительской особи. В этом случае новый организм может развиться из одной клетки, а возникшие потомки по своим наследственным признакам идентичны материнскому организму. Бесполое размножение позволяет быстро увеличить число особей. Различают два вида бесполого размножения:
- спорообразование – возникновение двух или более особей в результате прорастания специализированных клеток – спор (малярийный плазмодий, хлорелла, хламидомонада, мхи, папоротники и т. д.);
- вегетативное размножение – возникновение двух или более особей в результате деления, или почкования материнского организма.
Бесполое размножение у одноклеточных (бактерии, протисты, грибы): происходит путем деления материнской клетки пополам.
Почкование заключается в том, что на материнской клетке первоначально образуется бугорок, содержащий ядро. Почка растет, достигает размеров материнской клетки и затем отделяется от нее (дрожжевые грибы, некоторые инфузории).
Шизогония – множественное деление клетки (малярийный плазмодий)……?
Спорогония – в результате мейотических делений образуются специализированные гаплоидные клетки спорозоиты (малярийный плазмодий).
Бесполое размножение у многоклеточных .
Почкование : почка состоит из эктодермы и энтодермы. Она растет, удлиняется и на переднем конце образуется ротовое отверстие с окружающими его щупальцами. Почкование завершается образованием маленького полипа, который может отделиться от материнского организма (гидра пресноводная). Множественное почкование без отделения от родительских особей (коралл красный) приводит к образованию колоний.
Фрагментация : из части тела регенерируется самостоятельная особь (морские звезды, многие черви, в частности, дождевой червь, молочная планария).
Вегетативное: размножение у растений вегетативными частями тела (черенки, усы, отводки, клубни, листья и. д.).
Спорообразование – широко распространено у споровых растений (водоросли, мхи, плауны, папоротники) и грибов. Споры покрыты плотной оболочкой, которая защищает клетки от неблагоприятных внешних воздействий.
Полиэмбриония : развитие нескольких особей из одного зародыша (характерно для броненосцев; у человека появление однояйцевых близнецов довольно редко).
Шизогония : бесполо размножение путем разделения тела на большое количество дочерних особей (фораминиферы, радиолярии).
Таким образом, при любых форма бесполого размножения все потомки имеют генотип, идентичный материнскому.
Бесполое размножение более выгодно для организмов, обитающих в относительно постоянных условиях. Оно приводит к увеличению численности особей данного вида, но не сопровождается повышением генетического разнообразия внутри вида. Поскольку основным клеточным механизмом бесполого размножения является митоз, особи дочернего поколения оказываются точными копиями родительского организма. Новые признаки, которые могут оказаться полезными при изменении условий среды, появляются только в результате мутаций.
Бесполое размножение считается более древней формой размножения, возникшей в процессе развития жизни ранее полового.
3. Половое размножение.
Половое размножение появилось более 3 млрд. лет назад и встречается во всех крупных группах ныне живущих организмов. Сущность полового размножения заключается в объединении генетической информации от двух особей одного вида – родителей – в наследственном материале потомка. Таким образом, наследственный материал каждой дочерней особи представляет собой уникальную комбинацию генетической информации родителей. Образующиеся в процессе полового размножения организмы отличаются друг от друга по генотипу, признакам, свойствам, характеру приспособленности к условиям питания. Поскольку в норме рекомбинация каждой пары генов осуществляется в каждом поколении, то приспособительные комбинации генов возникают гораздо чаще за счет рекомбинаций, чем за счет относительно редких мутаций. Разнообразие генотипов особей, составляющих вид, обеспечивает возможность более успешного и быстрого приспособления вида к меняющимся условиям питания, освоения новых экологических ниш.
Биологическое значение полового размножения :
1. Самовоспроизведение особей.
2. Обеспечение биологического разнообразия видов, их адаптационных возможностей и эволюционных перспектив. Это позволяет считать половое размножение более прогрессивным, чем бесполое.
В основе полового размножения лежит половое процесс , т. е. обмен генетическим материалом между особями одного вида (конъюгация ) или объединение его (копуляция ).
Типичный половой процесс протекает у бактерий инфузорий и некоторых других организмов. Рассмотрим его особенности на примере инфузории-туфельки. Две инфузории подходят друг к другу брюшными сторонами, оболочка в месте их прикосновения растворяется и между ними образуется цитоплазматический мостик. Макронуклеусы при этом растворяются, и микронуклеусы делятся мейозом на четыре ядра, три из которых разрушаются, а четвертое вновь делится пополам митозом. В результате в каждой инфузории образуются два ядра – мужское (мигрирующее) и женское (стационарное). Затем происходит обмен мигрирующими ядрами с последующим слиянием стационарного и мигрирующего ядер, после чего особи расходятся. Вскоре в каждой из них ядро делится и впоследствии образуются микро- и макронуклеусы. Таким образом, при конъюгации число особей не увеличивается, а лишь возникают новые комбинации генетической информации.
Половое размножение, в отличие от бесполого, всегда происходит путем слияния двух специализированных половых клеток – яйцеклеток и сперматозоидов, образующихся в половых железах. Основное направление эволюции полового размножения – сингамия , т. е. оплодотворение, при котором обязательно слияние двух половых клеток, происходящих от разных особей. Такой тип размножения наилучшим способ обеспечивает генетическое разнообразие потомства.
Размножение – это способность организмов производить себе подобных особей того же вида. Существует два типа размножения: половое и бесполое. Древнейшим способом размножения на Земле было бесполое размножение. Бесполое размножение. Бесполое размножение происходит без образования специальных клеток, в нём участвует один организм, одна особь, при этом размножении образуются идентичные потомки. Единственным источником генетической изменчивости являются случайные мутации. Цитологической основой бесполого размножения является митоз. Молекулярной основой бесполого размножения является репликация ДНК. Бесполое размножение у различных живых организмов может происходить по-разному. Формы бесполого размножения: 1. Почкование – это форма бесполого размножения при которой новая особь образуется в виде выростов (почки) на теле родительской особи, а затем отделяется от неё и превращается в самостоятельную особь (гидра, дрожжи). 2. Фрагментация – это разделение особи на две или более частей, каждая из которых растёт и образуется отдельная особь (высшие растения, губка, дождевой червь). 3. Образование спор. Спора – это одноклеточная репродуктивная единица, состоящая из ядра и небольшого количества цитоплазмы под плотной оболочкой. Из споры образуется новая особь (низшие растения). 4. Деление. Бинарное деление клетки на две части. Ядро родительской особи один или несколько раз делится митозом, при этом образуется два или несколько дочерних ядер. Каждое из них окружается цитоплазмой и развивается в самостоятельный организм. 5. Шизогония – это множественное деление клетки. Сначала в клетке многократно делится ядро, затем вокруг каждого ядра обособляется участок цитоплазмы, который окружается плазматической мембраной. Затем происходит распад на отдельные клетки (малярийный плазмодий). 6. Вегетативное размножение. Осуществляется формирование дочернего организма из группы клеток материнского организма. У растений это размножение происходит за счёт вегетативных органов: корневищ, луковиц, клубней, усов. В результате бесполого размножения образуются генетически идентичные особи. Скорость размножения очень высокая и в постоянных условиях организма быстро захватывают экологическую нишу. Половое размножение. Появилось половое размножение более 3 млрд. лет назад. Сущность полового размножения в перекомбинации генетического материала родительских особей. В результате дочерние особи становятся более разнообразными, и естественный отбор выбирает из них наиболее приспособленные. При половом размножении потомство получается в результате слияния гаплоидных клеток – гамет. При оплодотворении образуется зигота. Из которой развивается новый организм. Оплодотворение – это процесс слияния сперматозоида с яйцеклеткой с последующим слиянием их ядер и образованием диплоидной зиготы. Биологическое значение этого процесса состоит в том, что при слиянии мужских и женских гамет образуется новый организм, несущий признак обоих родительских организмов. Гаметы гаплоидны, они содержат половинный набор хромосом и образуются в результате мейоза. Одной из модификаций полового размножения является партеногенез. Партеногенез – это процесс, при котором женская гамета развивается в новую особь без оплодотворения (встречается у животных (пчёлы) и растений). Преимущество в том, что увеличивается скорость размножения.
Эволюция размножения стала важной движущей силой общей эволюции царства растений и привела к возникновению новых специализированных органов. Бесполое размножение - наиболее древний и, несомненно, изначальный способ размножения. Оно представлено у всех групп растений, включая покрытосеменные
Наиболее примитивный способ бесполого размножения - вегетативное. Его наблюдают на самых ранних этапах развития жизни - у доклеточных. У многих предъядерных (например, у бактерий) это и сейчас единственный способ размножения. Каждая крупная естественная группа растений нередко обладает свойственными только ей формами вегетативного размножения.
Так, у сине-зеленых водорослей это гормогонии, у лишайников - изидии и соредии, у грибов - хламидоспоры и почкование, у некоторых моховидных- выводковые тельца. Наиболее разнообразны формы вегетативного размножения у высших растений, особенно у покрытосеменных. Велико разнообразие спор собственно бесполого размножения. В процессе эволюции они утратили способность воспроизводить новую особь, сходную с материнской (спорофит), а дают начало половому поколению (гаметофиту). Половое размножение возникло на очень ранних этапах эволюции, но время его появления точно не установлено.
У некоторых современных примитивных групп растений половой процесс неизвестен. Не вызывает сомнений, что такие природные группы, как сине-зеленые водоросли и большинство бактерий, никогда не имели полового размножения. Другие, например некоторые грибы, возможно, утратили его. Половой процесс утрачен и у некоторых высокоорганизованных растений, например у покрытосеменных (партеногенез). В настоящее время наука еще не располагает достаточными данными для воспроизведения полной картины эволюции полового процесса у растений.
Твердо установлено, что у растений, стоящих на более низкой ступени эволюции, половой процесс протекает в более примитивной форме, чем у растений, стоящих на более высокой ступени эволюции. Один из показателей уровня эволюции - специализация гамет. Наиболее примитивен изогамный половой процесс. Более высокий уровень - гетерогамия.
Наиболее специализированным половым процессом считают оогамию. Мужская гамета теряет подвижность, приспособившись к наземным условиям жизни, и доставляется к яйцеклетке пыльцевой трубкой. Таким образом, у семенных растений половой процесс не связан с наличием водной среды, это - приспособление к сухопутному образу жизни. Половое размножение развивалось не изолированно от бесполого, а во взаимной связи с ним. Каждая природная группа растений в жизненном цикле имеет свою закономерность смены бесполого и полового размножения. У многих низших растений смена ядерных фаз не выражена, спорофит почти полностью выпадает из жизненного цикла (диплоидна лишь зигота).
В жизненном цикле некоторых низших растений и всех высших существует чередование спорофита и гаметофита. Степень их морфологической разнородности и преобладания в жизненном цикле служит важным показателем эволюционного уровня изучаемой группы растений. На основании работ В. Гофмейстера (1849, 1851) установлена важная общая закономерность: в процессе эволюции спорофит приобретает всё большее развитие и самостоятельность; гаметофит, напротив, все более редуцируется, полностью теряет свою самостоятельность и всецело зависит от спорофита, что и наблюдают у наиболее высокоорганизованных семенных растений - голосеменных и покрытосеменных. В. Гофмейстер, изучая жизненные циклы различных природных групп растений, показал, что между высшими споровыми и покрытосеменными нет такого большого разрыва, как предполагали ранее.
12,13. Овогенез. Характеристика основных периодов. Строение женских половых гонад.
Сперматогенез. Характеристика основных периодов. Строение мужских половых гонад.
Гаметогенез - процесс образования яйцеклеток (овогенез) и сперматозоидов (сперматогенез) -подразделяется наряд стадий (рис. 5.4).
В стадии размножения диплоидные клетки, из которых образуются гаметы, называют сперматогониями и овогониями. Эти клетки осуществляют серию последовательных митотических делений, в результате чего их количество существенно возрастает. Сперматогонии размножаются на протяжении всего периода половой зрелости мужской особи. Размножение овогоний приурочено главным образом к периоду эмбриогенеза. У человека в женском организме этот процесс наиболее интенсивно протекает в яичниках между 2-м и 5-м месяцами внутриутробного развития. К 7-му месяцу большая часть овоцитов входит в профазу I мейоза.
Так как способом размножения клеток-предшественниц женских и мужских гамет является митоз, то овогоний и сперматогонии, как и все соматические клетки, характеризуются диплоидностью. В ходе митотического цикла их хромосомы имеют либо однонитчатую (после митоза и до завершения синтетического периода интерфазы), либо двунитчатую (постсинтетический период, профаза и метафаза митоза) структуру в зависимости от количества биспиралей ДНК. Если в одинарном, гаплоидном наборе число хромосом обозначить как п, а количество ДНК - как с, то генетическая формула клеток в стадии размножения соответствует 2п2с до S-периода и 2n4c после него.
Рис. 5.4. Схема гаметогенеза:
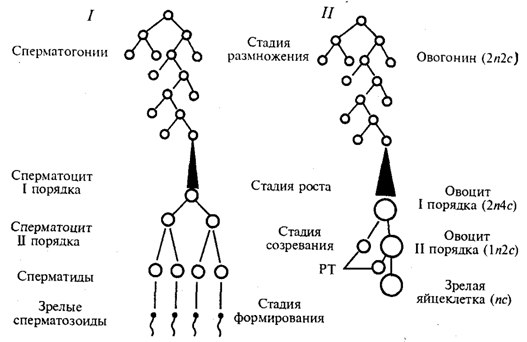
1 - сперматогенез, 2 - овогенез, n - количество хромосомных наборов,с - количество ДНК, РТ - редукционные тельца
На стадии роста происходит увеличение клеточных размеров и превращение мужских и женских половых клеток в сперматоциты и овоциты I порядка, причем последние достигают больших размеров, чем первые. Одна часть накапливаемых веществ представляет собой питательный материал (желток в овоцитах), другая - связана с последующими делениями. Важным событием этого периода является репликация ДНК при сохранении неизменным числа хромосом. Последние приобретают двунитчатую структуру, а генетическая формула сперматоцитов и овоцитов I порядка приобретает вид 2n4с.
Основными событиями стадии созревания являются два последовательных деления: редукционное и эквационное, которые вместе составляют мейоз (см. разд. 5.3.2). После первого деления образуются сперматоциты и овоциты II порядка (формула n2с), а после второго - сперматиды и зрелая яйцеклетка (пс).
В результате делений на стадии созревания каждый сперматоцит I порядка дает четыре сперматиды, тогда как каждый овоцит I порядка - одну полноценную яйцеклетку и редукционные тельца, которые в размножении не участвуют. Благодаря этому в женской гамете концентрируется максимальное количество питательного материала - желтка.
Процесс сперматогенеза завершается стадией формирования, или спермиогенеза. Ядра сперматид уплотняются вследствие сверхспирализации хромосом, которые становятся функционально инертными. Пластинчатый комплекс перемещается к одному из полюсов ядра, образуя акросомный аппарат, играющий большую роль в оплодотворении. Центриоли занимают место у противоположного полюса ядра, причем от одной из них отрастает жгутик, у основания которого в виде спирального чехлика концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, так что головка зрелого сперматозоида практически ее лишена.
МЕЙОЗ (деления созревания, период созревания), этап в образовании половых клеток; состоит из двух последовательных делений исходной диплоидной клетки (содержат два набора хромосом – 2n) и формирования четырёх гаплоидных половых клеток, или гамет (содержат по одному набору хромосом – n). Уменьшение (редукция) числа хромосом (2nn) происходит за счёт того, что на два деления приходится лишь одно удвоение (репликация ) хромосомного материала. При оплодотворении гаплоидные гаметы – яйцеклетка и сперматозоид – сливаются и диплоидное число хромосом, характерное для каждого вида, восстанавливается (n + n2n). В главных чертах мейоз протекает сходно у разных групп организмов и у особей женского и мужского пола. Два следующих друг за другом деления первичной половой клетки обозначаются как мейоз I и мейоз II. Подобно делению соматических клеток – митозу, и мейоз I, и мейоз II состоят из четырёх основных стадий – профазы, метафазы, анафазы и телофазы. Вступающая в мейоз клетка диплоидна, а каждая хромосома содержит удвоенное количество ДНК. В первом мейотическом делении особенно сложна и длительна профаза I (у человека она занимает 22,5 сут). На этой стадии гомологичные хромосомы соединяются (конъюгируют) в пары – биваленты. В каждой хромосоме бивалента различимы в микроскопе две продольные половины – хроматиды , т. е. бивалент представляет собой четвёрку (тетраду) хроматид. В профазе I происходит генетически значимое событие – обмен гомологичными (содержащими одни и те же гены) участками несестринских хроматид, или кроссинговер . В анафазе I биваленты разъединяются и гомологичные хромосомы расходятся к противоположным полюсам клетки, причем, в отличие от анафазы митоза, каждая хромосома сохраняет две хроматиды. В результате число хромосом уменьшилось вдвое, но удвоенным остаётся и количество ДНК, представленное двумя хроматидами. Важная особенность расхождения хромосом заключается в том, что любая, отцовская или материнская, хромосома из гомологичной пары может отойти к любому из полюсов независимо от того, как расходятся хромосомы других пар. Это означает, что число возможных сочетаний хромосом в дочерних клетках обычно очень велико: 2n , где n – число хромосомных пар (у человека – 223 ). Так происходит ещё одно перемешивание родительского генетического материала – рекомбинация хромосом. После мейоза I обычно сразу или после короткой интерфазы, во время которой удвоение хромосом не происходит, следует мейоз II. Это деление аналогично митозу с той разницей, что делятся гаплоидные клетки. В анафа-зе II сестринские хроматиды разделяются и, став хромосомами, расходятся к полюсам. Число хромосом и количество ДНК приходят в соответствие, и мейоз II завершается образованием четырёх гаплоидных гамет, каждая из которых несёт уникальный генетический материал. У самок, однако, лишь одна из четырёх гамет – яйцеклетка, способная к оплодотворению. Мейоз – один из ключевых биологических процессов. Его значение состоит в поддержании в поколениях постоянства хромосомных наборов (кариотипов ), т. е. в обеспечении наследственности , и в создании новых сочетаний отцовских и материнских генов, т. е. в обеспечении генотипической изменчивости .
Бесполое размножение с помощью одноклеточных спор свойственно и различным грибам и водорослям. Споры в этом случае образуются путем митоза (митоспоры), причем иногда (особенно у грибов) в огромных количествах; при прорастании они воспроизводят материнский организм. Некоторые грибы, например злостный вредитель растений фитофтора, образуют подвижные, снабженные жгутиками споры, называемые зооспорами или бродяжками. Проплавав в капельках влаги некоторое время, такая бродяжка «успокаивается», теряет жгутики, покрывается плотной оболочкой и затем, в благоприятных условиях, прорастает. Помимо митоспор, у многих из указанных организмов, а также у всех высших растений формируются споры и иного рода, а именно мейоспоры, образующиеся путем мейоза. Они содержат гаплоидный набор хромосом и дают начало поколению, обычно не похожему на материнское и размножающемуся половым путем. Таким образом, образование мейоспор связано с чередованием поколений – бесполого (дающего споры) и полового.
2.3. Вегетативное размножение
Другой вариант бесполого размножения осуществляется путем отделения от организма его части, состоящей из большего или меньшего числа клеток. Из них развивается взрослый организм. Примером может служить почкование у губок и кишечнополостных или размножение растений побегами, черенками, луковицами или клубнями. Такая форма бесполого размножения обычно называется вегетативным размножением. В своей основе оно аналогично процессу регенерации. Вегетативное размножение играет важную роль в практике растениеводства. Так, может случиться, что высеянное растение (например, яблоня) обладает некой удачной комбинацией признаков. У семян данного растения эта удачная комбинация почти наверняка будет нарушена, так как семена образуются в результате полового размножения, а оно связано с рекомбинацией генов. Поэтому при разведении яблонь обычно используют вегетативное размножение - отводками, черенками или прививками почек на другие деревья.
Бесполое размножение, воспроизводящее идентичные исходному организму особи, не способствует появлению организмов с новыми вариантами признаков, а тем самым ограничивает возможность приспособления видов к новым для них условиям среды. Средством преодоления этой ограниченности стал переход к половому размножению.
3. Половое размножение
Принципиальное отличие полового размножения от бесполого состоит в том, что в нем участвуют обычно два родительских организма, признаки которых перекомбинируются у потомства. Половое размножение свойственно всем эукариотам, но преобладает оно у животных и высших растений.
Переход к этому типу размножения имел огромное значение для эволюции жизни на Земле. Половое размножение создает бесконечное разнообразие особей, в том числе и таких, которые успешно адаптируются к изменчивым внешним условиям, «завоевывают мир», распространяясь в новые места обитания, и оставляют потомство, передавая ему свой наследственный материал. Потомки же двух успешных родительских особей могут оказаться обладателями еще более удачной комбинации наследственных признаков, и соответственно они разовьют успех родителей. Особи с неудачной комбинацией признаков будут элиминированы естественным отбором. Таким образом, половое размножение создает богатый материал для естественного отбора и эволюции. Любопытно и другое: само возникновение особи как индивидуальности, неделимого и смертного существа, является результатом перехода к половому размножению. При бесполом размножении клетка бесконечно делится, повторяя саму себя: она потенциально бессмертна, но особью может быть названа только условно, так как не отличима от неопределенного множества дочерних клеток. При половом размножении, напротив, все потомки различаются между собой и отличаются от родителей, а те с течением времени умирают, унося с собой свойственные им неповторимые особенности. Американский зоолог Р.Хегнер, обсуждая простейших, выразил это таким образом: «Они приобрели очередное новшество – пол; цена этого приобретения – неминуемая естественная гибель... Не велика ли эта цена?» Подчеркнем однако, что одновременно открылись возможности для развития и совершенствования, и они привели к появлению разнообразных живых форм, не сопоставимых по уровню организации с теми организмами, которые остановились на бесполом размножении.
3.1. Половое размножение животных
Переход к половому размножению связан с появлением специализированных половых клеток – мужских и женских гамет, в результате слияния которых (оплодотворения) образуется зигота – клетка, из которой развивается новый организм, обладающий новой комбинацией исходных генетических признаков.
Половое размножение впервые появилось у простейших, но переход к нему не был связан с немедленной утратой способности к репродукции бесполым путем: ряд животных сохранили ее, обычно чередуя бесполое размножение с половым. Такое чередование поколений наблюдается у некоторых простейших, кишечнополостных и оболочников.
3.2. Гаметы и гонады
Основой образования гамет (гаметогенеза) служит мейоз – клеточное деление с уменьшением вдвое числа хромосом, вследствие чего гаметы, в отличие от всех других клеток организма, гаплоидны. Слияние гамет восстанавливает число хромосом в зиготе до диплоидного. Последующее деление зиготы происходит путем митоза. Отметим, что у всех многоклеточных организмов деление всех клеток тела, кроме половых, происходит путем митоза. Следовательно, бесполое размножение клеток посредством деления надвое сохранилось в эволюции как основной механизм роста и развития организма, но не его репродукции.
У многих простейших половое размножение происходит с участием морфологически одинаковых мужских и женских гамет (у фораминифер, например, они представлены очень мелкими клетками, образующимися в гаплоидной родительской клетке в цикле чередования поколений). Такое явление называется изогамией. Она свойственна только одноклеточным.
Однако уже у некоторых простейших, например споровиков, и у всех многоклеточных организмов произошла дифференциация гамет: они стали различаться по форме и функции – возникла гетерогамия, т.е. разделение половых клеток на яйца (женские гаметы) и сперматозоиды (мужские гаметы).
Большинству животных свойственна т.н. оогамия: крупная неподвижная яйцеклетка (яйцо) и мелкий подвижный сперматозоид, за счет активных движений которого происходит его контакт с яйцом, ведущий к оплодотворению.
У губок и некоторых ресничных червей половые клетки рассеяны в теле и выводятся через разрывы стенки тела или через ротовое отверстие, но у многих плоских червей (а в зачаточной форме – и у гидры) появились гонады – специальные железы, продуцирующие гаметы. Мужские гонады – это семенники, женские – яичники. Правда, у таких гермафродитных животных, как брюхоногие моллюски, мужские и женские половые клетки созревают в одной и той же гонаде, но обычно в разное время, так что гонада функционирует то как семенник, то как яичник, и самооплодотворения не происходит. У других гермафродитных животных, например плоских червей или пиявок, одна особь содержит и яичники, и семенники; однако даже в случае одновременного созревания яиц и сперматозоидов животное избегает самооплодотворения и обычно спаривается с другой особью (исключение составляют, например, солитеры (цепни), одиночно живущие в кишечнике). Гермафродитизм наиболее распространен у червей и моллюсков и редко встречается у более высокоорганизованных форм – иглокожих, членистоногих и позвоночных; с другой стороны, он довольно редок и у таких древнейших многоклеточных, как кишечнополостные и в частности медузы.
Уже у некоторых червей и моллюсков в дополнение к гонадам сформировались половые протоки – семяпроводы и яйцеводы. Гонады и половые протоки составляют основные функциональные части внутренних половых органов, и они имеются у всех более высокоорганизованных животных.
3.3. Осеменение
Половые органы обеспечивают продукцию и выделение половых клеток, а тем самым – осеменение, т.е. сближение яиц и сперматозоидов двух особей. Процесс осеменения предшествует оплодотворению – слиянию гамет. Различают два способа осеменения (и соответственно оплодотворения): наружное и внутреннее. При наружном осеменении яйца и сперматозоиды выделяются в воду, где сперматозоиды, активно плавая, могут соединиться с яйцом и произвести оплодотворение. Понятно, что этот способ может быть свойствен только водным (или, как земноводные, сохранившим связь с водной средой) животным, и действительно, он наблюдается у большинства из них. Наружное осеменение обычно не связано со сложным устройством половой системы, хотя у некоторых животных развиваются дополнительные приспособления, например, для сцепления двух особей во время выделения ими половых продуктов.
Бóльшую независимость от внешних факторов (в частности, от водной среды) и более экономную продукцию гамет обеспечивает другой способ осеменения – внутреннее, при котором сперматозоиды вводятся непосредственно в женские половые пути. Известен также вариант внутреннего осеменения с помощью сперматофоров – капсул, наполненных сперматозоидами. Такое осеменение называют иногда наружно-внутренним.
У саламандры, например, самка захватывает выделенный самцом сперматофор своей клоакой, куда открываются половые протоки; самцы многих паукообразных с помощью своих клешневидных хелицер (первой пары головных конечностей) переносят сперматофор прямо в половое отверстие самки; самец головоногих моллюсков захватывает сперматофор особым видоизмененным щупальцем и переносит его в мантийную полость самки. Но в любом случае оплодотворение происходит внутри тела самки, обычно в яйцеводах. Оплодотворенные яйца откладываются во внешнюю среду (у большинства видов) или же развиваются внутриутробно. Внутреннее осеменение свойственно ряду водных животных и всем наземным. Оно появилось уже на очень ранней ступени эволюции, а именно у плоских червей.
3.4. Усложнение половой системы
Переход к внутреннему осеменению и оплодотворению сопровождался усложнением половой системы и формированием дополнительных половых органов. Так, образовались железы, например выделяющие жидкость, в которой находятся сперматозоиды и которая необходима им для движения, или – у самок – формирующие наружную оболочку яиц. У плоских червей и ряда других животных, особенно у насекомых, развились семяприемники для хранения поступающей при осеменении спермы. Поскольку сперматозоиды могут длительно сохранять в них жизнеспособность, наличие семяприемников делает оплодотворение менее зависимым от встречи партнеров: многие насекомые успешно размножаются, спариваясь единственный раз в жизни. Соответственно и время между спариванием и откладкой яиц может варьировать в широких пределах.
У самок ряда насекомых (стрекоз, цикад, кузнечиков, наездников и др.) образовался такой дополнительный половой орган, как яйцеклад, служащий для откладки яиц в ячейки, землю либо в ткани растений или животных.
Возникли также копулятивные (совокупительные) органы как приспособление для внутреннего осеменения. У разных групп животных они формировались разным путем: у многих из них – из нижнего отдела полового протока, но, например, у ракообразных – путем видоизменения одной пары ножек, у мух и других двукрылых насекомых – из конечных сегментов брюшка, у живородящих рыб – из выростов плавников. Однако у ряда животных, например многих птиц, специальные копулятивные органы отсутствуют.
Если у некоторых яйцекладущих совершенствовался аппарат для откладки яиц, то у животных, перешедших к живорождению, прежде всего у млекопитающих, произошли иные изменения половой системы; самое значительное из них – преобразование среднего отдела яйцевода в матку, где развивается зародыш.
3.5. Спаривание
Одним из условий успешного размножения служит одновременное созревание гамет у мужских и женских особей. Некоторые животные способны размножаться круглый год, но у многих, особенно у обитателей средних и высоких широт, размножение сезонное. В этом случае наступление периода размножения зависит от внешних факторов: длины светового дня, температуры воздуха, наличия пищи и т.д. Действие этих факторов на репродуктивную систему, как правило, не прямое, а опосредованное – чаще всего гормонами, регулирующими функциональную активность гонад и/или уровень метаболизма. Так, у позвоночных с сезонным размножением изменение освещенности влияет на секрецию гормонов гипофиза, «включающих» функцию гонад, а тем самым и определяющих сроки размножения.
Однако этих физиологических механизмов может быть недостаточно для обеспечения спаривания. В действие часто вступает половой отбор наиболее сильных и приспособленных особей, обычно самцов, способных привлечь самку и отстоять свое право на размножение. Турнирные бои между самцами, ухаживание перед спариванием, охрана своей территории для размножения, так же как, по-видимому, и брачный наряд самцов, – все это средства достижения успеха в размножении самых жизнеспособных особей. Половое поведение достигает большой сложности у высокоорганизованных животных с их развитой нейроэндокринной системой.
Большинство животных не образует постоянных пар, и проблема поиска партнера для спаривания возникает у них регулярно. Однако среди птиц и млекопитающих встречаются моногамные виды, т.е. образующие прочные пары (например, волки, лебеди, попугаи). Известны примеры и полигамии; так, морские котики, тюлени, некоторые другие млекопитающие и птицы создают устойчивую группу из одного – более сильного, чем его конкуренты, – самца и целого гарема самок.
4. Способы воспроизведения потомства
Разные группы животных выработали не только разные способы оплодотворения; у них по-разному появляется на свет потомство. В зависимости от того, как это происходит, различают три способа размножения.
Яйцерождение. Подавляющее большинство видов животных откладывают яйца, из которых выводится молодь. Таких животных называют яйцеродящими или яйцекладущими. К ним относятся почти все морские беспозвоночные, насекомые, многие рыбы, земноводные, пресмыкающиеся, птицы и однопроходные млекопитающие.
Живорождение. У живородящих животных оплодотворенное яйцо развивается в теле самки, получая от нее питание до самого рождения детеныша на свет. К живородящим относятся все млекопитающие за исключением однопроходных – утконоса и ехидны. Живорождение встречается и в других группах, например у некоторых пресмыкающихся и у более примитивных животных.
Яйцеживорождение. Существует и промежуточная форма воспроизведения потомства: яйцо развивается, продолжая оставаться в теле самки, но питание зародыша обеспечивается желтком яйца, а не организмом матери. Яйцеживорождение свойственно некоторым акулам и другим рыбам, ряду земноводных, многим ящерицам и змеям.
Стратегии размножения. С этими способами размножения связаны и разные его стратегии. На одном полюсе – стратегия экономного размножения, характеризующегося медленным воспроизведением малочисленного потомства и заботливым его выращиванием (выкармливанием, уходом, обереганием, научением); на другом – расточительное, избыточное размножение с производством очень большого количества яиц и воспроизведением многочисленного потомства при отсутствии заботы о нем. Если в первом случае вероятность выживания потомства весьма значительна, то во втором шансы на сохранение яиц и выживание каждого отдельного потомка крайне малы, так что только интенсивное размножение может компенсировать высокий процент гибели яиц и молоди на всех стадиях развития. Первая стратегия в целом свойственна высокоорганизованным животным – млекопитающим и птицам. Однако в пределах этих групп основная стратегия может быть выражена в разной степени.
Например, у приматов она проявляется в наибольшей мере: они длительно вынашивают плод и производят на свет обычно по одному, еще очень беспомощному, детенышу, которого долго вскармливают и растят. С другой стороны, мыши, крысы или кролики могут плодиться несколько раз в год, рождая каждый раз до десятка детенышей, которые быстро приобретают самостоятельность. В результате такого интенсивного размножения повышается вероятность гибели потомков из-за недостатка пищи или – в связи с быстрым ростом популяции – из-за распространения болезней и размножения хищников. Таким образом, сравнивая грызунов с приматами, стратегию их размножения следует признать неэкономной. Тем не менее расточительность сил при размножении грызунов не сопоставима с тем, что наблюдается у различных видов яйцекладущих, например рыб, многие из которых выметывают сотни тысяч и миллионы икринок.
Многим животным свойственна забота о сохранности яиц: одни откладывают их в ил, землю и разные укромные места, другие (в частности, некоторые ракообразные и офиуры, среди рыб морская игла и морской конек, среди земноводных жабы-повитухи и пипы) носят яйца на себе, и количество яиц в этом случае много меньше, чем при выметывании их в воду. Еще дальше в этой стратегии пошли яйцеживородящие.
Своеобразную стратегию размножения избрали общественные насекомые, например муравьи и общественные пчелы. Они строят гнезда, оберегают яйца и обеспечивают питанием личинок, но оставляют функцию размножения только одной (у пчел) или нескольким (у муравьев) самкам в сообществе. Размножающаяся самка, называемая маткой или царицей, откладывает многочисленные яйца. Самцы появляются лишь на короткое время и после спаривания погибают.
Партеногенез. Яйца некоторых организмов способны развиваться без оплодотворения, т.е. без участия сперматозоида. Такой процесс однополого размножения называют партеногенезом, или девственным размножением. Его рассматривают как редуцированную форму полового размножения.
Примеры естественного партеногенеза у млекопитающих неизвестны; они изредка встречаются у низших позвоночных и весьма обычны у беспозвоночных, особенно у насекомых. Существует два типа партеногенеза: облигатный (т.е. обязательный) и факультативный. Первый свойствен видам, у которых самцов либо нет совсем, либо они редки и не способны функционировать. К таким видам относятся некоторые тли, палочники, сверчки, бабочки; популяции без самцов изредка встречаются у рыб, например у серебряного карася. При факультативном партеногенезе яйца могут развиваться как партеногенетически, так и в результате оплодотворения, причем партеногенетическое размножение может преобладать в условиях, когда слишком редки контакты разнополых особей, например на границе ареала распространения вида.
Известен также циклический партеногенез, при котором размножение с участием обоих полов чередуется с партеногенетическим. Например, многие виды тлей дают несколько партеногенетических поколений в течение короткого теплого периода лета, а на зиму откладывают оплодотворенные яйца, которые покрыты плотной оболочкой и способны перезимовывать; весной из них выходят только самки, но осенью появляется поколение с некоторым количеством самцов – и цикл возобновляется. Аналогичным образом размножаются и некоторые другие виды с высокой сезонной смертностью, например коловратки. Циклический партеногенез наблюдается также у видов с личиночным размножением; при этом оплодотворенные яйца откладывают обычно только зрелые особи, а у личинок они развиваются партеногенетически.
Заключение
В заключении, хотелось бы подчеркнуть тот факт, что размножение, в направлении от бесполых форм к половым, от изогамии к анизогамии, (Гаметы) от участия всех клеток в размножении к разделению клеток на соматические и половые, от наружного оплодотворения к внутреннему с внутриутробным развитием и заботой о потомстве – все виды в большей своей части схожи лишь тем, что обеспечивают заботу о своем потомстве, такую необходимую, при размножении.
Литература
1. Большая Советская Энциклопедия. Размножение
2. Гартман М., Общая биология, пер. с нем., М. - Л., 1936
3. Гормональная регуляция размножения у млекопитающих. М., 1987
4. Докинз Р. Эгоистичный ген. М., 1993
5. Мэйнард Смит Дж. Эволюция полового размножения. М., 1981
6. Мясоедов С. В., Явления размножения и пола в органическом мире, Томск, 1935
7. Справочник школьника по биологии для 6-9 классов.
Гормональная регуляция размножения у млекопитающих. М., 1987
Мэйнард Смит Дж. Эволюция полового размножения. М., 1981
Н.А. Монтеверде Малая современная энциклопедия
Большая советская энциклопедия. Размножение
Введение 3
1. Типы размножения 4
1.1 Бесполое размножение 4
1.2 Половое размножение 6
2. Индивидуальное развитие организмов 12
2.1 Эмбриональный период развития 12
2.2 Постэмбриональный период развития 16
2.3 Общие закономерности развития. Биогенетический закон 18
Заключение 21
Список литературы 22
Введение
Способность размножаться, т.е. производить новое поколение особей того же вида, - одна из основных особенностей живых организмов. В процессе размножения происходит передача генетического материала от родительского поколения следующему поколению, что обеспечивает воспроизведение признаков не только данного вида, но конкретных родительских особей. Для вида смысл размножения состоит в замещении тех его представителей, которые гибнут, что обеспечивает непрерывность существования вида; кроме того, при подходящих условиях размножение позволяет увеличить общую численность вида.
Каждая новая особь, прежде чем достигнуть стадии, на которой она будет способна к размножению, должна пройти ряд стадий роста и развития. Некоторые особи погибают, не достигнув репродуктивной стадии (или половозрелости) в результате уничтожения хищниками, болезней и разного рода случайных событий; поэтому вид может сохраниться лишь при условии, что каждое поколение будет производить больше потомков, чем было родительских особей, принимавших участие в размножении. Численность популяций колеблется в зависимости от баланса между размножением и вымиранием особей. Существует ряд различных стратегий размножения, каждая из которых имеет определенные преимущества и недостатки; все они будут описаны в этом реферате.
1. Типы размножения
Известны различные формы размножения, но все они могут быть объединены в два типа: половое и бесполое.
Половым размножением называют смену поколений и развитие организмов из специализированных – половых – клеток, образуются в половых железах. При этом новый организм развивается в результате слияния двух половых клеток, образованных разными родителями. Однако у беспозвоночных животных нередко сперматозоиды и яйцеклетки формируются в теле одного организма. Такое явление - обоеполость - называют гермафродитизмом. Цветковые растения также бывают обоеполыми. У большинства видов покрытосеменных (цветковых) растений обоеполый цветок включает и тычинки, образующие мужские половые клетки - спермин, и пестики, содержащие яйцеклетки. Примерно у четвертой части видов мужскиe (тычиночные) и женские (пестичные) цветки развиваются независимо, т.е. у них цветки однополые. Примером может служить конопля. У некоторых растений - кукурузы, березы - и мужские и женские цветки возникают на одной особи.
У
некоторых видов
животных и
растений наблюдается
развитие
неоплодотворенной
яйцеклетки.
Такое размножение
называют девственным
или партеногенетическим.
Бесполое размножение характеризуется тем, что новая особь развивается из неполовых, соматических (телесных) клеток.
1.1 Бесполое размножение
При бесполом размножении новый организм может возникнуть из одной клетки или из нескольких неполовых (соматических) клеток материнской особи. В бесполом размножении участвует только одна родительская особь. Поскольку клетки, дающие начало дочерним организмам, возникают в результате митоза, то все потомки окажутся сходными по наследственным признакам с материнской особью.
Рис. 1. Размножение эвглены зеленой
Многие простейшие (амебы, эвглена зеленая и др.), одноклеточные водоросли (хламидомонада) размножаются путем митотического деления клетки (рис. 1). Другим одноклеточным - некоторым низшим грибам, водорослям (хлорелла), животным, например возбудителю малярии - малярийному плазмодию, свойственно спорообразование. При этом клетка распадается на большое число особей, равное количеству ядер, заранее образованных в родительской клетке в результате многократного деления ее ядра. Многоклеточные организмы также способны к спорообразованию: это мхи, высшие грибы, многоклеточные водоросли, папоротникообразные и некоторые другие.
Как у одноклеточных, так и у многоклеточных организмов способом бесполого размножения служит также почкование. Например, у дрожжевых грибов и некоторых инфузорий (сосущие инфузории) при почковании на материнской клетке первоначально образуется небольшой бугорок, содержащий ядро, - почка. Она растет, достигает размеров, близких к размерам материнского организма, и затем отделяется, переходя к самостоятельному существованию. У многоклеточных (пресноводная гидра) почка состоит из группы клеток обоих слоев стенки тела. Почка растет, удлиняется, на переднем ее конце появляется ротовое отверстие, окруженное щупальцами. Почкование завершается образованием маленькой гидры, которая затем отделяется от материнского организма.
У многоклеточных животных бесполое размножение осуществляется таким же путем (медузы, кольчатые черви, плоские черви, иглокожие). Из каждой такой части развивается полноценная особь.
У растений широко распространено вегетативное размножение, т.е. частями тела - черенками, усами, клубнями. Так, картофель размножается видоизмененными подземными частями стебля - клубнями. У жасмина, ивы легко укореняются побеги - черенки. С помощью черенков размножают виноград, смородину, крыжовник.
Длинные ползучие стебли земляники - усы - образуют почки, которые, укореняясь, дают начало новому растению. Немногие растения, например бегония, могут размножаться листовыми черенками (листовая пластинка и черешок). На нижней стороне листа, в местах разветвления крупных жилок, возникают корни, на верхней – почки, а затем побеги.
Для вегетативного размножения используют также корень. В садоводстве с помощью черенков из боковых корней размножают малину, вишню, сливу, розу. С помощью корневых клубней размножаются георгины. Видоизменение подземной части стебля - корневище - также образует новые растения. Например, осот с помощью корневища может дать более тысячи новых особей на 1 м2 почвы.
1.2 Половое размножение
Половое размножение имеет очень большие эволюционные преимущества по сравнению с бесполым. Это обусловлено тем, что генотип потомков возникает путем объединения генов, принадлежащих обоим родителям. В результате повышаются возможности организмов в приспособлении к условиям окружающей среды. Так как новые комбинации осуществляются в каждом поколении, то приспособленными к новым условиям существования может оказаться гораздо большее количество особей, чем при бесполом размножении. Появление новых комбинаций генов обеспечивает более успешное и быстрое приспособление вида к меняющимся условиям обитания.
Таким образом, сущность полового размножения заключается в объединении в наследственном материале потомка генетической информации из двух разных источников - родителей.
В половых железах развиваются половые клетки: мужские - сперматозоиды, женские - яйцеклетки (или яйца). В первом случае их развитие называют сперматогенезом, во втором - овогенезом (от лат. ово - яйцо).
В процессе образования половых клеток выделяют ряд стадий. Первая стадия - период размножения, в котором первичные половые клетки делятся путем митоза, в результате чего увеличивается их количество.
Вторая стадия - период роста. У незрелых мужских гамет он выражен не резко. Их размеры увеличиваются незначительно. Напротив, будущие яйцеклетки - овоциты - увеличиваются в размерах иногда в сотни, а чаще в тысячи и даже миллионы раз. Рост овоцитов осуществляется за счет веществ, образуемых другими клетками организма. Так, у рыб, амфибий и в большей степени у рептилий и птиц основную массу яйца составляет желток. Он синтезируется в печени, в особой растворимой форме переносится кровью в яичник, проникает в растущие овоциты и откладывается там в виде желточных пластинок. Кроме того, в самой будущей половой клетке синтезируются многочисленные белки и большое количество разнообразных РНК: транспортных, рибосомных и информационных. Желток - совокупность питательных веществ (жиров, белков, углеводов, витаминов и др.), необходимых для питания развивающегося зародыша, а РНК обеспечивает синтез белков на ранней стадии развития, когда собственная бедственная информация еще не используется.
Следующая стадия - период созревания, или мейоз, - представлена на рисунке 2. Клетки, вступающие в период созревания, содержит диплоидный набор хромосом и уже удвоенное количество ДНК.
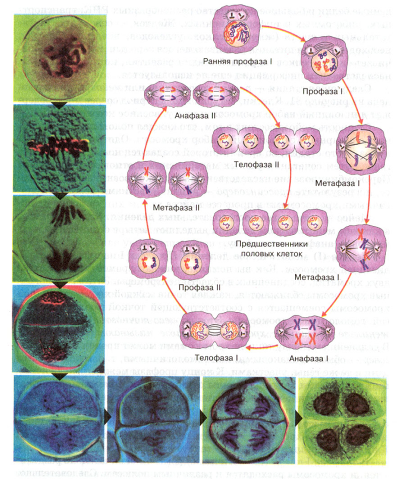
Рис. 2. Созревание половых клеток (мейоз)
Сущность мейоза состоит в том, что каждая половая клетка получает одинарный, гаплоидный, набор хромосом. Однако вместе с тем мейоз - это стадия, во время которой создаются новые комбинации генов путем сочетания разных материнских и отцовских хромосом, рекомбинирование наследственных задатков возникает, кроме того в результате кроссинговера - обмена участками между гомологичными хромосомами в процессе мейоза.
Мейоз включает два последовательных деления. Как и в митозе, в каждом мейотическом делении выделяют четыре стадии: профазу, метафазу, анафазу и телофазу.
Первое (I) мейотическое деление. Профаза I начинается спирализацией хромосом. Как вы помните, каждая хромосома состоит из двух хроматид, соединенных в области центромеры. Затем гомологические хромосомы сближаются, каждая точка каждой хроматиды одной хромосомы совмещается с соответствующей точкой хроматиды другой, гомологичной хромосомы. Этот процесс точного и тесного сближения гомологичных хромосом в мейозе называют конъюгацией. В дальнейшем между такими хромосомами может произойти кроссинговер - обмен одинаковыми, или гомологичными, т. е. содержащими одни и те же гены, участками. К концу профазы между гомологичными хромосомами возникают силы отталкивания. Вначале они проявляются в области центромер, а затем в других участках.
В метафазе I спирализация хромосом максимальна. Конъюгированные хромосомы располагаются по экватору, причем центромеры гомологичных хромосом обращены к разным полюсам клетки. К ним прикрепляются нити веретена деления.
В анафазе I плечи гомологичных хромосом окончательно разделяются, и хромосомы расходятся к различным полюсам. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна. Число хромосом уменьшается в два раза, хромосомный набор становится гаплоидным. Однако каждая хромосома состоит из двух хроматид, т. е. по-прежнему содержит удвоенное количество ДНК.
В телофазе I на непродолжительное время образуется ядерная оболочка. Во время интерфазы между первым и вторым делениями мейоза редупликации ДНК не происходит. Клетки, образовавшиеся в результате первого деления созревания, различаются по составу отцовских и материнских хромосом и, следовательно, по набору генов.
Например, все клетки человека, в том числе первичные половые клетки, содержат 46 хромосом. Из них 23 получены от отца и 23 - от матери. При образовании половых клеток после первого мейотического деления в сперматоциты и овоциты также попадает по 23 хромосомы. Однако вследствие случайности расхождения отцовских и материнских хромосом в анафазе I образующиеся клетки получают самые разнообразные комбинации родительских хромосом. Например, в одной из них может оказаться 3 отцовских и 20 материнских хромосом, в другой - 10 отцовских и 13 материнских, в третьей - 20 отцовских и 3 материнских и т. д. Число возможных комбинаций очень велико. Если учесть еще обмен гомологичными участками хромосом в профазе первого деления мейоза, то вполне очевидно, что каждая образующаяся половая клетка генетически уникальна, так как несет свой неповторимый набор генов.
Следовательно, мейоз - основа комбинативной генотипической изменчивости.
Второе (II) мейотическое деление. Второе деление мейоза в общем протекает так же, как обычное митотическое деление, с той лишь разницей, что делящаяся клетка гаплоидна. В анафазе II центромеры, соединяющие сестринские хроматиды в каждой хромосоме, делятся, и хроматиды, как и в митозе, с этого момента становятся самостоятельными хромосомами. С завершением телофазы II заканчивается и весь процесс мейоза: из исходной первичной половой клетки образовались четыре гаплоидные клетки.
У особей мужского пола все они преобразуются в гаметы - сперматозоиды. У особей женского пола вследствие неравномерного мейоза лишь из одной клетки получается жизнеспособное яйцо. Три другие дочерние клетки гораздо мельче, они превращаются в так называемые направительные, или редукционные, тельца, вскоре погибающие. Образование только одной яйцеклетки и гибель трех генетически полноценных направительных телец с биологической точки зрения обусловлено необходимостью сохранения в одной клетке всех запасных питательных веществ, которые понадобятся для развития будущего зародыша.
Период формирования состоит в приобретении клетками определенной формы и размеров, соответствующих их функции.
Женские половые клетки в процессе созревания покрываются оболочками и готовы к оплодотворению непосредственно после завершения мейоза. Во многих случаях, например у пресмыкающихся, птиц и млекопитающих, за счет деятельности клеток, окружающих яйцеклетку, вокруг нее возникает ряд дополнительных оболочек. Их функция заключается в защите яйцеклетки и развивающегося зародыша от внешних неблагоприятных воздействий. Сперматозоиды могут иметь различные размеры и форму.
Функция сперматозоидов состоит в доставке в яйцеклетку генетической информации и стимуляции ее развития. Сформированный сперматозоид содержит митохондрии, аппарат Гольджи, выделяющий ферменты, растворяющие мембрану яйца при оплодотворении, т. е. при слиянии сперматозоида и яйцеклетки. Возникающая при этом диплоидная клетка носит название зиготы.
2. Индивидуальное развитие организмов
Индивидуальным развитием, или онтогенезом, называют весь период жизни особи - с момента слияния сперматозоида с яйцом и образования зиготы до гибели организма. Онтогенез делится на два периода: 1) эмбриональный - от образования зиготы до рождения или выхода из яйцевых оболочек; 2) постэмбриональный - от выхода из яйцевых оболочек или рождения до смерти организма.
Наука, изучающая закономерности индивидуального развития организмов на стадии зародыша, называется эмбриологией (от греч. эмбрион - зародыш).
2.1 Эмбриональный период развития
У большинства многоклеточных животных, независимо от сложности их организации, стадии эмбрионального развития, которые проходит зародыш, едины. В эмбриональном периоде выделяют три основных этапа: дробление, гаструляцию и первичный органогенез.
Дробление. Развитие организма начинается со стадии одной клетки. Оплодотворенное яйцо - это клетка и одновременно уже организм на самой ранней стадии его развития. В результате многократных делений одноклеточный организм превращается в многоклеточный. Возникшее при оплодотворении путем слияния сперматозоида и яйцеклетки диплоидное ядро через несколько минут начинает делиться, вместе с ним делится и цитоплазма. Образующиеся клетки с каждым делением уменьшаются в размерах, поэтому процесс деления носит название дробления. В период дробления накапливается клеточный материал для дальнейшего развития. Завершается дробление образованием многоклеточного зародыша - бластулы. Бластула имеет полость, наполненную жидкостью, так называемую первичную полость тела.
В тех случаях, когда в цитоплазме яйцеклетки желтка мало (как у ланцетника) или относительно немного (как у лягушки), дробление бывает полным, т. е. яйцеклетка делится целиком.
Иначе протекает период дробления у птиц. Свободная от желтка цитоплазма составляет всего 1% от общего объема яйцеклетки курицы; вся остальная цитоплазма яйцеклетки, а следовательно и зигота, заполнена массивом желтка. Если присмотреться к куриному яйцу, на одном из его полюсов непосредственно на желтке можно увидеть маленькое пятнышко - бластулу, или зародышевый диск, образовавшийся в результате дробления свободного от желтка участка цитоплазмы, содержащего ядро. В таких случаях дробление называют неполным. Неполное дробление свойственно и некоторым рыбам и рептилиям.
Во всех случаях - и у ланцетника, и у амфибий, и у птиц, а также у других животных - общий объем клеток на стадии бластулы не превышает объема зиготы. Другими словами, митотическое деление зиготы не сопровождается ростом образовавшихся дочерних клеток до объема материнской, и размеры их в результате ряда последовательных делений прогрессивно уменьшаются. Эта особенность митотического деления клеток в ходе дробления наблюдается при развитии оплодотворенных яиц у всех животных.
Некоторые другие черты дробления также свойственны различным видам животных. Например, все клетки в бластуле имеют диплоидный набор хромосом, одинаковы по строению и отличаются друг от друга главным образом количеством содержащегося в них желтка. Такие клетки, лишенные признаков специализации для выполнения определенных функций, называют неспециализированными (или недифференцированными) клетками. Другая особенность дробления - чрезвычайно короткий митотический цикл бластомеров по сравнению с клетками взрослого организма. Во время очень короткой интерфазы происходит только удвоение ДНК.
Гаструляция. Бластула, как правило, состоящая из большого числа бластомеров (например, у ланцетника из 3000 клеток), в процессе развития переходит в новую стадию, которую называют гаструлой (от греч. гастер - желудок). Зародыш на этой стадии состоит из отчетливо различимых пластов клеток - так называемых зародышевых листков: наружного, или эктодермы (от греч. эктос - находящийся снаружи), и внутреннего, или энтодермы (от греч. энтос - находящийся внутри). Совокупность процессов, приводящих к образованию гаструлы, называют гаструляцией.
У ланцетника гаструляция осуществляется путем впячивания одного из полюсов бластулы внутрь, по направлению к другому, у других животных - либо путем расслоения стенки бластулы, либо путем обрастания массивного вегетативного полюса мелкими клетками анимального полюса.
У многоклеточных животных, кроме кишечнополостных, параллельно с гаструляцией или, как у ланцетника, вслед за ней возникает и третий зародышевый листок - мезодерма (от греч. мезос - находящийся посередине), которая представляет собой совокупность клеточных элементов, расположенных между экто- и энтодермой в первичной полости тела - бластоцеле. С появлением мезодермы зародыш становится трехслойным.
Таким образом, сущность процесса гаструляции заключается перемещении клеточных масс. Клетки зародыша практически делятся и не растут. Однако на этой стадии начинается использование генетической информации клеток зародыша, появляются первые признаки дифференцировки.
Дифференцировка, или дифференцирование, - это процесс ее возникновения и нарастания структурных и функциональных различий между отдельными клетками и частями зародыша. С морфологической точки зрения дифференцирование выражается в том, что образуются несколько сотен типов клеток специфического строения, отличающихся друг от друга. Из неспециализированных клеток бластулы постепенно возникают клетки эпителия кожи, эпителия кишечника, легких, появляются нервные, мышечные клетки и т.д. С биохимической точки зрения специализация клеток заключается в способности синтезировать определенные белки, свойственные только данному типу клеток. Лимфоциты синтезируют защитные белки - антитела, мышечные клетки - сократительный белок миозин. Каждый тип клеток образует «свои», свойственные только ему белки. Биохимическая специализация клеток обеспечивается избирательной - дифференциальной активностью генов, т. е. в клетках разных зародышевых листков - зачатков определенных органов и систем - начинают функционировать разные группы генов.
У разных видов животных одни и те же зародышевые листки дают начало одним и тем же органам и тканям. Это значит, что они гомологичны. Так, из клеток наружного зародышевого листка - эктодермы - у членистоногих, хордовых, в том числе у рыб, амфибий, рептилий, птиц и млекопитающих, формируются кожные покровы и их производные, а также нервная система и органы чувств. Гомология зародышевых листков подавляющего большинства животных - одно из доказательств единства животного мира.
Органогенез. После завершения гаструляции у зародыша образуется комплекс осевых органов: нервная трубка, хорда, кишечная трубка. У ланцетника осевые органы формируются следующим образом: эктодерма на спинной стороне зародыша прогибается по средней линии, превращаясь в желобок, а эктодерма, расположенная справа и слева от него, начинает нарастать на его края. Желобок - зачаток нервной системы - погружается под эктодерму, и края его смыкаются. Образуется нервная трубка. Вся остальная эктодерма - зачаток кожного эпителия.
Спинная часть энтодермы, располагающаяся непосредственно под нервным зачатком, обособляется от остальной энтодермы и сворачивается в плотный тяж - хорду. Из оставшейся части энтодермы развиваются мезодерма и эпителий кишечника. Дальнейшая дифференцировка клеток зародыша приводит к возникновению многочисленных производных зародышевых листков - органов и тканей. В процессе специализации клеток, входящих в состав зародышевых листков, из эктодермы образуются нервная система, органы чувств, эпителий кожи, эмаль зубов; из энтодермы - эпителий кишки, пищеварительные железы - печень и поджелудочная железа, эпителий жабр и легких; из мезодермы - мышечная ткань, соединительная ткань, в том числе рыхлая соединительная ткань, хрящевая и костная ткани, кровь и лимфа, а также кровеносная система, почки, половые железы.
2.2 Постэмбриональный период развития
В момент рождения или выхода организма из яйцевых оболочек заканчивается эмбриональный и начинается постэмбриональный период развития. Постэмбриональное развитие может быть прямым или сопровождается превращением (метаморфозом).
При прямом развитии (у пресмыкающихся, птиц, млекопитающих) из яйцевых оболочек или из тела матери выходит организм небольших размеров, но с уже заложенными всеми основными органами, свойственными взрослому животному. Постэмбриональное развитие в этом случае сводится в основном к росту и половому созреванию.
При развитии с метаморфозом из яйца выходит личинка, обычно устроенная проще взрослого животного, со специальными личиночными органами, во взрослом состоянии отсутствующими. Личинка питается, растет, и со временем личиночные органы заменяются органами, свойственными взрослым особям. Следовательно, при метаморфозе разрушаются личиночные органы и возникают органы, присущие взрослым животным.
Разберем несколько примеров непрямого постэмбрионального развития. Личинка асцидий (тип Хордовые, подтип Личиночно-хордовые) обладает всеми основными признаками хордовых животных: хордой, нервной трубкой, жаберными щелями в глотке. Она свободно плавает, затем прикрепляется к какой-либо твердой поверхности на дне моря, где и совершается метаморфоз: у нее исчезают хвост, хорда, мышцы, а нервная трубка распадается на отдельные клетки, большая часть которых фагоцитируется. От нервной системы личинки остается лишь группа клеток, дающая начало нервному узлу. Строение тела взрослой асцидий, ведущей прикрепленный образ жизни, нисколько не напоминает обычные черты организации хордовых животных. Только знание особенностей онтогенеза позволяет определить систематическое положение асцидий: строение личинки указывает на происхождение их от хордовых животных, которые вели свободный образ жизни. В процессе метаморфоза асцидий переходят к сидячему образу жизни, в связи с чем упрощается их организация.
Личиночная форма амфибий - головастик, для которого характерны жаберные щели, боковая линия, двухкамерное сердце, один круг кровообращения. В процессе метаморфоза, происходящего под влиянием гормона щитовидной железы, рассасывается хвост, появляются конечности, исчезает боковая линия, развиваются легкие и второй круг кровообращения. Обращает на себя внимание сходство ряда черт строения головастиков и рыб (боковая линия, строение сердца и кровеносной системы, жаберные щели).
Примером метаморфоза может служить также развитие насекомых. Гусеницы бабочек или личинки стрекоз резко отличаются по строению, образу жизни и среде обитания от взрослых животных и напоминают своих предков - кольчатых червей.
Постэмбриональный период развития имеет разную продолжительность. Например, поденки в личиночном состоянии живут 2-3 года, а в половозрелом - от 2-3 часов до 2-3 суток в зависимости от видовой принадлежности. В большинстве же случаев постэмбриональный период более продолжителен. У человека он включает стадию полового созревания, стадию зрелости и стадию старости.
У млекопитающих
и человека
наблюдается
известная
зависимость
продолжительности
жизни от длительности
полового созревания
и беременности.
Обычно продолжительность
жизни превышает
дорепродуктивный
период онтогенеза
в 5-8 раз.
Постэмбриональное развитие сопровождается ростом. Различают рост неопределенный, продолжающийся в течение всей жизни, и определенный, ограниченный каким-то сроком. Неопределенный рост наблюдается у древесных форм растений, некоторых моллюсков, из позвоночных - у рыб, крыс.
У многих животных рост прекращается вскоре после достижения половой зрелости. У человека рост заканчивается к 20-25 годам.
2.3 Общие закономерности развития. Биогенетический закон
Все многоклеточные организмы развиваются из оплодотворенного яйца. Развитие зародышей у животных, относящихся к одному типу, во многом сходно. У всех хордовых животных в эмбриональном периоде закладывается осевой скелет - хорда, возникает нервная трубка, в переднем отделе глотки образуются жаберные щели. План строения хордовых животных также одинаков. На ранних стадиях развития зародыши позвоночных очень похожи (рис. 3). Эти факты подтверждают справедливость сформулированного К. Бэром закона зародышевого сходства: «Эмбрионы обнаруживают, уже начиная с самых ранних стадий, известное общее сходство в пределах типа». Сходство зародышей разных систематических групп свидетельствует об общности их происхождения. В дальнейшем в строении зародышей проявляются признаки класса, рода, вида и, наконец, признаки, характерные для данной особи. Расхождение признаков зародышей в процессе развития называется эмбриональной дивергенцией и отражает эволюцию той или иной систематической группы животных, историю развития данного вида.

Рис. 3. Зародышевое сходство у позвоночных: 1 – однопроходные (ехидна), 2 – сумчатые (кенгуру), 3 – парнокопытные (олень), 4 – хищные (кошка), 5 – приматы (мартышка), 6 - человек
Большое
сходство зародышей
на ранних стадиях
развития и
по
явление
различий на
более поздних
стадиях имеют
свое объяснение.
Организм
подвержен
изменчивости
на протяжении
всего развития.
Мутационный
процесс затрагивает
гены, обусловливающие
особенности
строения и
обмена веществ
у самых молодых
эмбрионов. Но
возникающие
у них структуры
(древние признаки,
свойственные
далеким предкам)
играют весьма
важную роль
в процессах
дальнейшего
развития. Как
указывалось,
зачаток хорды
индуцирует
образование
нервной трубки,
а его утрата
приводит к
прекращению
развития. Поэтому
изменения на
ранних стадиях
обычно приводят
к недоразвитию
и гибели особи.
Напротив, изменения
на поздних
стадиях, затрагивая
менее значительные
признаки, могут
быть благоприятны
для организма
и в таких случаях
подхватываются
естественным
отбором.
Появление в эмбриональном периоде развития современных животных признаков, свойственных их далеким предкам, отражает эволюционные преобразования в строении органов.
В своем развитии организм проходит одноклеточную стадию (стадия зиготы), что может рассматриваться как повторение филогенетической стадии первобытной амебы. У всех позвоночных, включая высших их представителей, закладывается хорда, которая далее замещается позвоночником, а у их предков, если судить по ланцетнику, хорда оставалась на всю жизнь. В ходе эмбрионального развития птиц и млекопитающих, включая человека, появляются жаберные щели в глотке и соответствующие им перегородки. Факт закладки частей жаберного аппарата у зародышей наземных позвоночных объясняется их происхождением от рыбообразных предков, дышащих жабрами. Строение сердца человеческого зародыша в ранний период формирования напоминает строение этого органа у рыб: оно с одним предсердием и одним желудочком. У беззубых китов в эмбриональном периоде появляются зубы. Зубы эти не прорезываются, они разрушаются и рассасываются.
Приведенные здесь и многие другие примеры указывают на глубокую связь между индивидуальным развитием организмов и их историческим развитием. Эта связь нашла свое выражение в биогенетическом законе, сформулированном Ф. Мюллером и Э. Геккелем в XIX в.: онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития) вида, к которому эта особь относится.
Заключение
Завершая работу над рефератом можно прийти к выводу, что способность к размножению, или самовоспроизведению, – одна из важнейших характеристик органической природы. Размножение – свойство, присущее всем без исключения живым организмам, от бактерий до млекопитающих.
Существование любого вида животных и растений, бактерий и грибов, преемственность между родительскими особями и их потомством поддерживаются только благодаря размножению. Тесно связано с самовоспроизведением и другое свойство живых организмов – развитие. Оно также присуще всему живому на Земле: и мельчайшим одноклеточным организмам, и многоклеточным растениям и животным.
Список литературы
Боген Г. Современная биология. - М.: Мир, 1970.
Грин Н., Стаут У., Тейлор Д. Биология: в 3-х т. Т. 3: пер. с англ./под ред. Р. Сопера. - М.: Мир, 1990.
Мамонтов С.Г. Биология. Общие закономерности. – М.: Дрофа, 2002.
От животных к человеку. – М.: Наука, 1971.
Слюсарев А.А. Биология с общей генетикой. - М.: Медицина, 1978.


