Клеточное строение корня. Анатомическое строение корня
По происхождению корни делятся на главные , боковые и придаточные. Главный корень семенных растений развивается из корешка зародыша семени. Стебель является продолжением корня, и вместе они составляют ось 1-го порядка. Место сочленения оси и семядольных листьев называют семядольным узлом. Участок, расположенный на границе главного корня и стебля, именуется корневой шейкой. Участок стебля от корневой шейки до первых зародышевых листьев (семядолей) называют подсемядольным коленом , или гипокотилем, а от семядолей до первых настоящих листьев - эпикотилем , или надсемядольным коленом . У двудольных и голосеменных растений от главного корня за счет меристематической активности перицикла отходят боковые корни 1-го порядка, которые дают начало боковым корням 2-го и 3-го порядка. Корневая система, образованная системой главного корня, называется стержневой, а с развитой системой боковых корней - ветвистой; таким образом, ветвистая корневая система является разновидностью стержневой. Чем больше боковых корней отходит от главного, тем больше площадь питания растения.
У большинства двудольных растений главный корень сохраняется всю жизнь, у однодольных растений главный корень не развивается, так как зародышевый корешок быстро отмирает, а от базальной части побега берут начало придаточные корни. Придаточные корни могут образовываться от листьев, стеблей, старых корней и даже от цветков по происхождению: А - система главного корня; Б, В - смешанная корневая система; Г - придаточная корневая система; 1 - главный корень; 2 - боковые корни; 3 - придаточные корни; 4 - основания побегов и иметь ответвления 1-го, 2-го порядка и т.д. Корневая система, образованная придаточными корнями, называется мочковатой (см. рис. 3.2). У многих двудольных корневищных растений главный корень часто отмирает и преобладает система придаточных корней, отходящих от корневища (лютик ползучий, сныть обыкновенная).
Рис. 3.2. Типы корневых систем: по форме:
А, Б - стержневая; В, Г - мочковатая;
по происхождению: А - система главного корня; Б, В - смешанная корневая система; Г - придаточная корневая система; 1 - главный корень; 2 - боковые корни; 3 - придаточные корни; 4 - основания побегов
По отношению к субстрату корни бывают следующих типов: земляные - развиваются в почве; водные - находятся в воде (у плавающих водных растений); воздушные, развивающиеся в воздушной среде (у растений, имеющих корни на стволах и листьях).
Зоны корня
В молодом корне различают 4 зоны: деления, растяжения, всасывания, проведения (см. рис. 3.3).
К зоне деления относят верхушку конуса нарастания (протяженность менее 1 мм), где происходит активное митотическое деление клеток. Верхушечная меристема откладывает наружу клетки корневого чехлика, а внутрь - ткани остальной части корня. Эта зона состоит из тонкостенных паренхимных клеток первичной меристемы, которые прикрыты корневым чехликом, выполняющим защитную функцию при продвижении корня между частичками почвы. От соприкосновения с почвой клетки чехлика постоянно разрушаются, образуя слизь, которая предохраняет зону деления при трении о
почву и продвижении корня вглубь. У большинства растений корневой чехлик восстанавливается за счет первичной меристемы, а у злаков - за счет особой меристемы калиптрогена.
По теории гистогенов (Ганштейн, 1868), у большинства покрытосеменных растений апикальные меристемы состоят из 3 гистогенных слоев, различающихся направлением деления клеток и имеющих по 1-4 инициальные клетки. Самый наружный слой - дерматоген - формирует протодерму, из которой образуются клетки корневого чехлика и ризодерма - первичная покровно-всасывающая ткань в зоне всасывания. Средний слой - периблема - дает начало всем тканям первичной коры. Третий слой инициалей формирует плером, из которого развиваются ткани центрального осевого цилиндра.
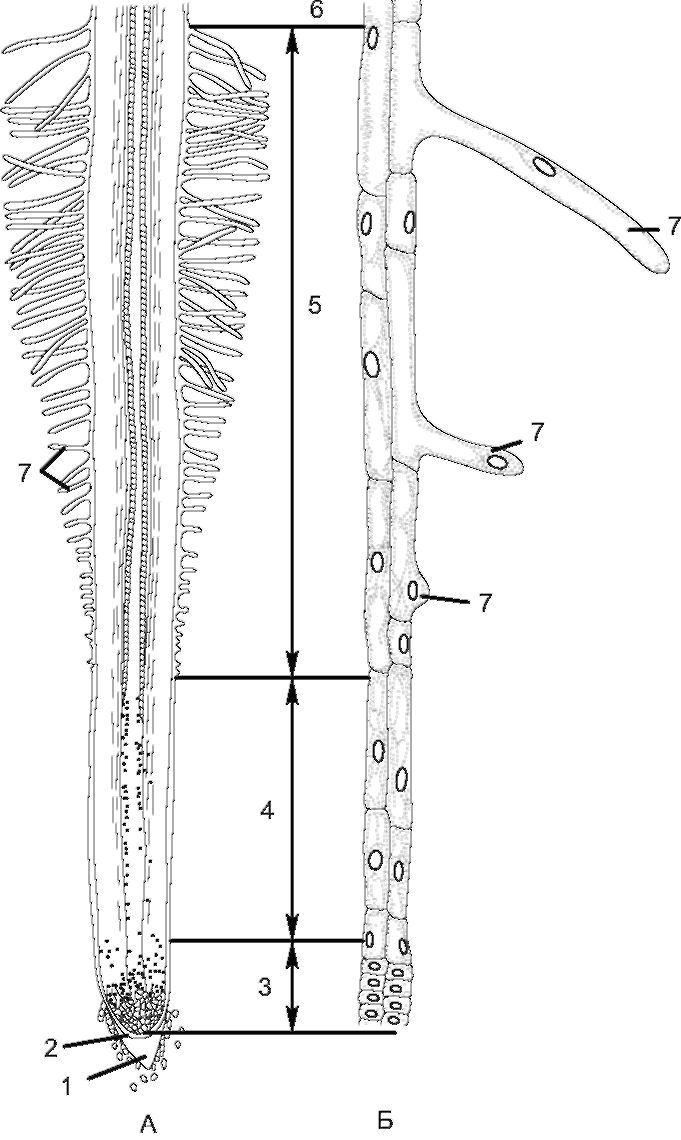
Рис. 3.3. Зоны корня (у проростка пшеницы):
А - схема строения корня; Б - периферические клетки отдельных зон при большом увеличении: 1 - корневой чехлик; 2 - калиптроген; 3 - зона деления; 4 - зона растяжения; 5 - зона всасывания; 6 - зона проведения;
7 - корневые волоски
В зоне растяжения клетки меристемы увеличиваются в размерах (вследствие оводнения), вытягиваются в длину, и деление клеток постепенно прекращается. Вследствие вытягивания клеток в продольном направлении происходят рост корня в длину и его продвижение в почве. Зону деления и зону растяжения с учетом сохранения в них меристематической активности можно объединить в одну - зону роста. Ее протяженность составляет несколько миллиметров. В зоне всасывания происходит становление первичной структуры корня.
Протяженность зоны всасывания - от нескольких миллиметров до нескольких сантиметров; она характеризуется наличием корневых волосков, являющихся выростами клеток ризодермы. При их формировании ядро перемещается в переднюю часть корневого волоска. Последние увеличивают всасывающую поверхность корня и обеспечивают активное всасывание воды и растворов солей, но они недолговечны (живут 10-20 дней). Новые корневые волоски образуются под зоной всасывания, а отмирают - над этой зоной. По мере роста растения зона всасывания постепенно перемещается, и растение имеет возможность поглощать минеральные вещества из разных слоев почвы.
Постепенно зона всасывания переходит в зону проведения (укрепления). Она тянется вплоть до корневой шейки и составляет по протяженности большую часть корня. В этой зоне идет интенсивное ветвление главного корня и появляются боковые корни. У двудольных растений в зоне проведения формируется вторичная структура корня.
Анатомия корня
Первичное строение корня (рис. 3, см. цв. вкл.). Строение корня в зоне всасывания называется первичным, потому что здесь происходит дифференциация тканей из первичной меристемы конуса нарастания. Первичное строение корня в зоне всасывания можно наблюдать у двудольных и однодольных растений, но у однодольных оно сохраняется на протяжении всей жизни растения. На поперечном срезе корня первичного строения выделяют 3 основные части: покровно-всасывающую ткань, первичную кору и центральный осевой цилиндр (рис. 3.4).
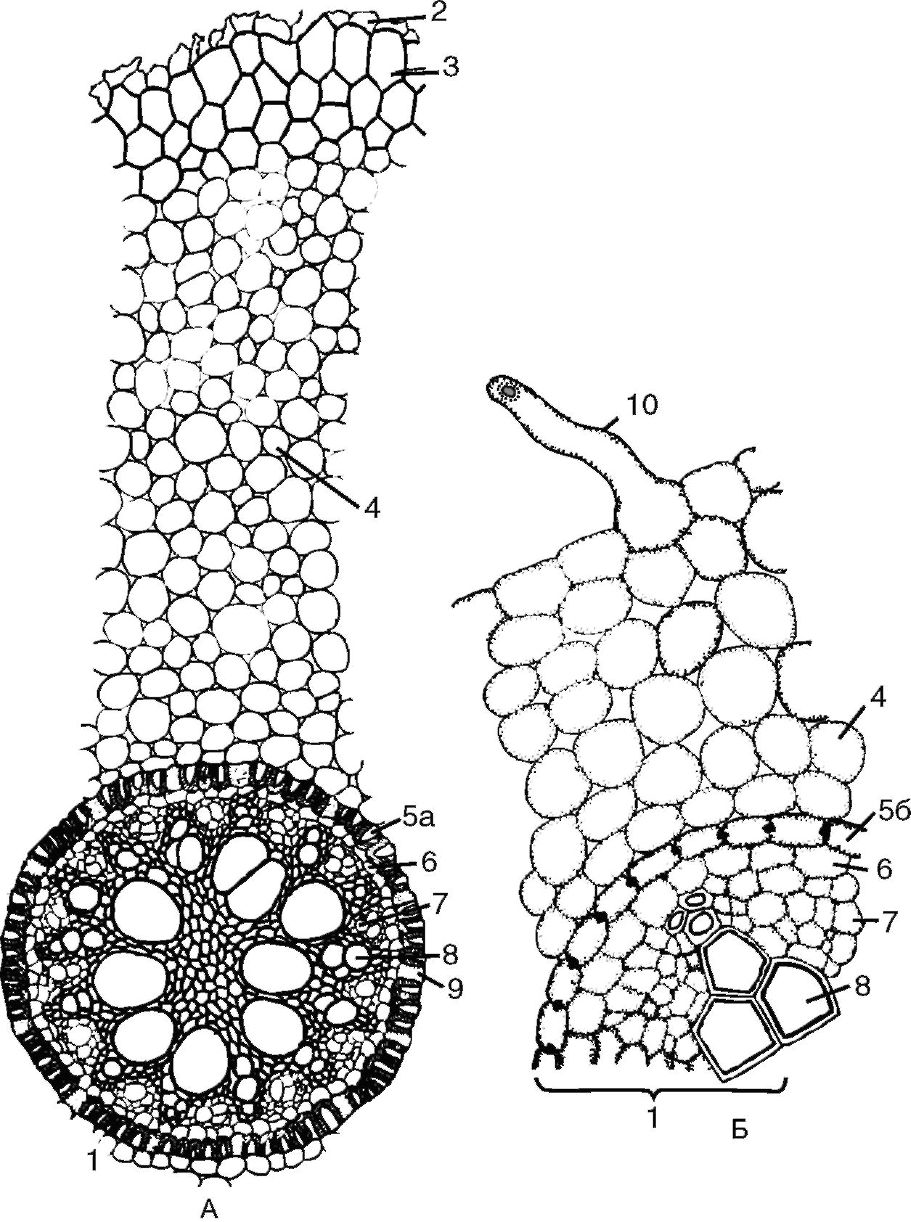
Рис. 3.4. Поперечный срез корня первичного строения:
А - первичное строение корня однодольного; Б - первичное строение корня двудольного: 1 - центральный (осевой) цилиндр; 2 - остатки эпиблемы; 3 - экзодерма; 4 - мезодерма; 5а - эндодерма с подковообразными утолщениями; 5б - эндодерма с поясками Каспари; 6 - перицикл; 7 - первичная флоэма; 8 - сосуды первичной ксилемы; 9 - пропускные клетки эндодермы; 10 - корневой волосок
Покровно-всасывающая ткань - ризодерма (эпиблема) выполняет как покровную функцию, так и функцию интенсивного всасывания воды и минеральных веществ из почвы. Клетки ризодермы живые, с тонкой целлюлозной стенкой. Из некоторых клеток ризодермы формируются корневые волоски; каждый из них представляет собой длинный вырост одной из клеток ризодермы, ядро же клетки обычно находится в кончике выроста. Корневой волосок содержит тонкий пристенный слой цитоплазмы, более плотный на верхушке волоска, а в центре - крупную вакуоль. Корневые волоски недолговечны и в зоне укрепления отмирают. Физиологически зона всасывания представляет собой очень важную часть корня. Клетки ризодермы поглощают водные растворы всей поверхностью наружных стенок. Развитие корневых волосков во много раз увеличивает поверхность поглощения. Протяженность зоны всасывания - от 1 до 1,5 см.
Со временем эпиблема может слущиваться, и тогда покровную функцию выполняет экзодерма, а после ее разрушения - слой кле- ток мезодермы и иногда мезодермы и перицикла, стенки которых опробковевают и одревесневают. Поэтому диаметр старых корней однодольных растений меньше, чем молодых.
Первичная кора корня развита более мощно, чем центральный осевой цилиндр. Она состоит из 3 слоев: экзодермы, мезодермы (рис. 4, см. цв. вкл.) (паренхима первичной коры) и эндодермы. Клетки экзодермы многоугольные по форме, плотно сомкнуты и расположены в несколько рядов. Клеточные стенки пропитаны суберином, т.е. опробковевают. Опробковение обеспечивает непроницаемость клеток для воды и газов. В экзодерме, обычно под корневыми волосками, сохраняются клетки с тонкими целлюлозными стенками - пропускные клетки, через которые проходят вода и минеральные вещества, поглощенные ризодермой. Обычно они располагаются напротив лучей ксилемы радиального пучка.
Под экзодермой находятся живые паренхимные клетки мезодермы . Это наиболее широкая часть первичной коры. Клетки мезодермы выполняют запасающую функцию, а также функцию проведения воды и растворенных в ней солей от корневых волосков в центральный осевой цилиндр.
Внутренний однорядный слой первичной коры представлен эндодермой. Клетки эндодермы плотно сомкнуты и почти квадратные в поперечном сечении. В зависимости от степени утолщения клеточной стенки различают 2 типа эндодермы - с поясками Каспари (на поперечном срезе они выглядят как пятна Каспари) и с подковообразными утолщениями стенок.
Эндодерма с поясками Каспари - это начальный этап формирования эндодермы, при котором утолщению подвергаются лишь ее радиальные стенки за счет отложения веществ, сходных по химическому составу с суберином и лигнином. У многих двудольных и голосеменных растений процесс дифференциации эндодермы поясками Каспари заканчивается. У эндодермы с подковообразными утолщениями образуется толстая вторичная клеточная стенка, пропитанная суберином, в дальнейшем она одревесневает. Неутолщенной остается только наружная клеточная стенка (рис. 3.5). Эндодерма с подковообразными утолщениями развивается чаще у однодольных растений (рис. 5, см. цв. вкл.).
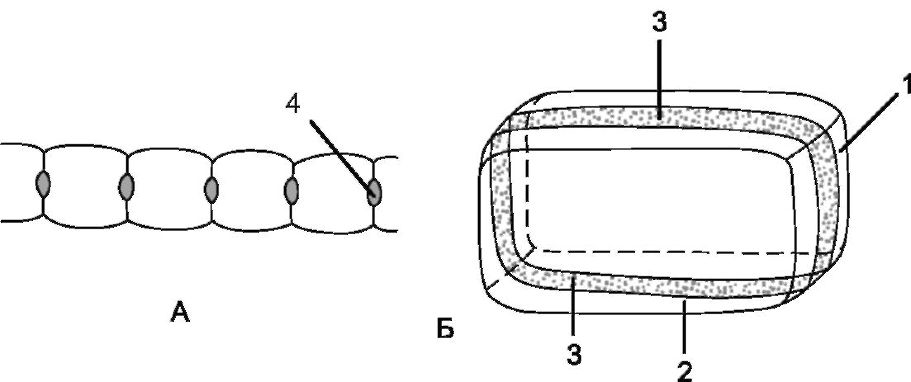
Рис. 3.5. Схема строения клетки эндодермы:
А - общий вид; Б - поперечный разрез клеток: 1 - поперечная стенка клетки; 2 - продольная радиальная стенка; 3 - поясок Каспари; 4 - пятна Каспари
Считается, что эндодерма выполняет функцию гидравлического барьера, способствуя продвижению минеральных веществ и воды из первичной коры в центральный осевой цилиндр и препятствуя их выходу обратно.
Центральный осевой цилиндр начинается с клеток перицикла, который обычно в молодых корнях состоит из живых тонкостенных паренхимных клеток, расположенных в один ряд (но может быть и многослойным - например, у грецкого ореха). Клетки перицикла дольше других тканей корня сохраняют свойства меристемы и спо- собность к новообразованиям. Из перицикла образуются боковые корни, поэтому его называют корнеродным слоем. Проводящая система корня представлена одним радиальным сосудисто-волокнистым пучком, в котором группы элементов первичной ксилемы чередуются с участками первичной флоэмы. У однодольных растений количество лучей первичной ксилемы - 6 и более, у двудольных - от 1 до 5. Корни в отличие от стеблей не имеют сердцевины, так как в центре корня располагаются лучи первичной ксилемы.
У однодольных и споровых архегониальных растений строение корня не претерпевает значительных изменений в течение всей жизни растения. У голосеменных и двудольных растений на границе зон всасывания и проведения происходит переход от первичного строения корня ко вторичному (табл. 3.1).
Анатомическое строение корня
Первичное строение корня. Оно характерно для молодых корней всех растений. У плаунов, хвощей, папоротников и однодольных растений оно сохраняется в течение всей жизни. Первичное строение возникает в результате дифференциации меристемы конуса нарастания. На поперечном срезе корня в зоне всасывания можно различить три части: эпиблему, первичную кору и центральный осевой цилиндр (стелу).
Эпиблема (ризодерма), или кожица – первичная покровная ткань корня. Состоит из одного ряда плотно сомкнутых клеток, имеющих выросты – корневые волоски.
Первичная кора состоит из живых тонкостенных клеток в периферической части корня. Представлена тремя четко отличающимися друг от друга слоями: экзодермой, мезодермой и эндодермой.
Экзодерма располагается непосредственно под эпиблемой, являясь наружной частью первичной коры. Ее клетки многоугольные, плотно сомкнутые, располагаются в один или несколько рядов. По мере отмирания корневых волосков экзодерма оказывается на поверхности корня и выполняет роль покровной ткани, при этом происходит утолщение и опробковение клеточных оболочек и отмирание содержимого клеток.
Мезодерма ,или основнаяпаренхима, располагается кнаружи от эндодермы. Состоит из рыхло расположенных клеток с системой межклетников, по которым идет интенсивный газообмен. Здесь происходит синтез и передвижение в другие ткани пластических веществ, накапливаются питательные вещества, располагается микориза.
Эндодерма – самый внутренний слой коры, непосредственно прилегающий к стеле. У двудольных растений он состоит из одного ряда клеток, имеющих утолщения на радиальных стенках, – пояски Каспари, непроницаемые для воды. У однодольных растений образуются подковообразные утолщения клеточных стенок. В эндодерме встречаются живые тонкостенные клетки, которые называют пропускными клетками . Эти клетки также имеют пояски Каспари. Клетки эндодермы контролируют поступление воды и растворенных в ней минеральных веществ из коры в центральный цилиндр и обратно.
Центральный цилиндр, или осевой цилиндр, или стела, занимает центральную часть корня. Наружный слой стелы, примыкающий к эндодерме, называется перициклом . Его клетки долго сохраняют способность к делению. Здесь закладываются боковые корешки. В центральной части осевого цилиндра находится сосудисто-волокнистый пучок. Для корней характерно чередование в стеле участков первичной ксилемы и первичной флоэмы . Ксилема образует звезду, а между ее лучами располагается флоэма. Количество лучей ксилемы различно – от двух до нескольких десятков, у двудольных – до пяти, у однодольных – более пяти. В самом центре цилиндра могут находиться элементы ксилемы, склеренхима или тонкостенная паренхима.
Вторичное строение корня. У двудольных и голосеменных растений первичное строение корня сохраняется недолго. Примерно через 10 дней после прорастания семян происходят изменения, в результате которых возникает вторичное строение корня.
Процесс вторичных изменений начинается с появления прослоек камбия под участками первичной флоэмы, внутрь от нее. Камбий возникает из слабо дифференцированной паренхимы центрального цилиндра. Внутрь он откладывает элементы вторичной ксилемы (древесины) , наружу – элементы вторичной флоэмы (луба). Сначала прослойки камбия разобщены, затем смыкаются, образуя сплошной слой. Это происходит благодаря делению клеток перицикла напротив лучей ксилемы. Камбиальные участки, возникшие из перицикла, образуют только паренхимные клетки серцевинных лучей, остальные клетки камбия образуют проводящие элементы – ксилему и флоэму. При делении клеток камбия исчезает радиальная симметрия, характерная для первичного строения корня.
В перицикле возникает и пробковый камбий (феллоген). Он откладывает наружу слои клеток вторичной покровной ткани – пробки. При этом первичная кора отмирает.
Корневые системы
 Корневая система –
это совокупность всех корней растения. В образовании корневой системы участвует главный корень, боковые и придаточные корни. По форме различают два основных типа корневых систем:
Корневая система –
это совокупность всех корней растения. В образовании корневой системы участвует главный корень, боковые и придаточные корни. По форме различают два основных типа корневых систем:
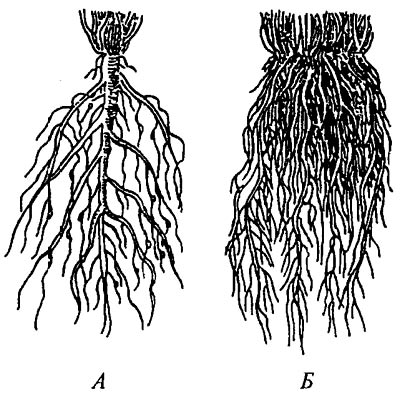
Стержневая корневая система(А) имеет хорошо выраженный главный корень. Она характерна для двудольных растений.
Мочковатая корневая система(Б) образована боковыми и придаточными корнями. Главный корень растет слабо и рано прекращает свой рост. Она типична для однодольных растений.
Почва
Для нормального роста и развития растений необходимы вода и питательные вещества, источником которых является почва. Почвой называют верхний корнеобитаемый, плодородный слой земной коры, в котором располагаются корни растений.
Любая почва состоит из трех главных компонентов:
Твердой фазы – мелкораздробленных простых и сложных минералов, органических веществ;
Жидкой фазы – почвенного раствора;
Газообразной фазы – почвенного воздуха.
Твердая фаза на 90% и более состоит из минералов и примерно на 10% из органических веществ – гумуса , образованного остатками растительного и животного происхождения. Количество гумуса в почве определяет ее плодородие. Содержание гумуса можно определить по цвету почвы: чем больше в почве гумуса, тем она темней.
Жидкая фаза представляет собой водный раствор различных минеральных солей, углекислоты, минеральных и органических кислот. Она служит непосредственным источником питательных веществ для растений.
Газообразная фаза служит источником кислорода для дыхания корней.
В основе классификации почв лежит размер частиц твердой фазы - от крупного гравия (свыше 2 мм в диаметре) до глины (диаметр частиц менее 0,002 мм). Различают каменистые, песчаные, суглинистые (50% песка, 25% пыли и 25% глины) и подзолистые почвы. Самыми благоприятными для произрастания растений являются черноземы - почвы, богатые перегноем. От механического состава почвы зависит ее влаго- и воздухоемкость.
Помимо гумуса почва содержит большое количество бактерий и грибов, принимающих участие в разложении органических остатков.
Удобрения
Для улучшения роста растений в почву вносят минеральные вещества и органические соединения - удобрения. Удобрением
называются органические или минеральные вещества, применяемые для улучшения условий питания растений.
Удобрения делят на две группы:
органические (навоз, торф, навозная жижа, птичий помет, фекалии, компосты, зеленое удобрение);
минеральные - азотные, фосфорные, калийные и другие промышленные удобрения, а их местных удобрений - зола.
Минеральные удобрения. Промышленные удобрения в зависимости от содержания основных питательных элементов делятся на:
простые , или односторонние - удобрения, содержащие в своем составе лишь один из трех важнейших питательных элементов (N, P или K) - азотные, фосфорные, калийные;
сложные
, или комбинированные
- удобрения, содержащие в своем составе два или три элемента: азотно-калийные, азотно-фосфорные, азотно-фосфорно-калийные (нитрофоски).
К важнейшим удобрениям относятся:
азотные - аммиачная селитра, карбамид (синтетическая мочевина), сульфат аммония, хлористый аммоний, натриевая селитра, кальциевая селитра (усиливают рост стеблей и листьев);
фосфорные - суперфосфат простой, суперфосфат двойной, фосфоритная мука, костяная мука (продлевают цветение, ускоряют созревание плодов);
калийные : хлористый калий, сульфат калия, карбонат калия, сернокислый калий (усиливают рост корней, луковиц, клубней);
комплексные удобрения : сульфатная нитрофоска, сернокислая нитрофоска, нитроаммофоска (N, P, K), аммофос, диаммофос (N, P), полифосфат калия, метафосфат калия (P, K).
Кроме N, P, K, требующихся растениям в значительных количествах, растениям необходимы и некоторые другие элементы, такие как бор, марганец, медь, молибден, цинк и другие. Эти элементы требуются не всем культурам и не на всех почвах в незначительных количествах. Они необходимы для получения высоких урожаев. Такие элементы получили название микроэлементов, а удобрения, их содержащие, - микроудобрениями . Микроудобрениями могут быть и отходы промышленности, и специально выпускаемые соединения.
Органические удобрения.
Достоинством органических удобрений является, прежде всего, их комплексность. Они соединяют в себе и минеральные соли, и органические вещества, образующие при разложении не только минеральные соединений, но и много углекислого газа, то есть улучшают и корневое и воздушное питание растений.
Одним из основных органических удобрений является навоз - отход животноводства, состоящий из выделений животных и подстилки. Органические вещества навоза становится доступным растениям лишь после минерализации. Этот процесс протекает медленно, поэтому в течение нескольких лет растения обеспечиваются необходимыми им веществами.
Вместе с навозом в почву вносится не только бесчисленное количество микробов, но и важный источник их пищи, что усиливает энергию биологических процессов в ней.
Применение навоза одновременно улучшает корневое и воздушное питание культур. Но это еще не все. Органические вещества навоза способствуют увеличению содержания в почве гумуса.
Применение удобрений.
Наиболее высокие прибавки урожая получают при правильном сочетании органических и минеральных удобрений, поскольку они взаимодополняют друг друга в целом ряде отношений.
Польза от совместного применения или сочетания органических и минеральных удобрений заключается в том что:
- органические удобрения действуют медленно, минеральные быстро; для растений создаются лучшие условия питания в том случае, когда внесены обе группы удобрений;
- органические вещества поглощают излишки минеральных удобрений и затем постепенно отдают их;
- органические удобрения доставляют пищу полезным микробам, которые превращают ее в необходимые растениям соли;
- органические удобрения улучшают структуру и свойства почвы.
Действие удобрений на растения зависит не только от их вида, состава, растворимости, но и от способа внесения в почву. Существует три способа применения удобрений:
· Основное удобрение вносят перед вспашкой и заделывают в почву (навоз, торф и не менее двух третей минеральных удобрений, предназначенных для культуры). Основное удобрение растения используют для питания в течение большей части вегетационного периода.
· Припосевное удобрение вносится в небольших количествах при посеве семян, посадке клубней, корней, рассады. Оно снабжает растения хорошо доступной пищей в начале роста, когда корневая система еще слаборазвита. Поэтому в качестве припосевного удобрения используют легко растворимые в воде и быстро усваиваемые растениями вещества.
· Подкормка
- внесение легкоусвояемых удобрений в сухом или растворенном виде во время роста растений. При подкормке обычно вносят вещества, наиболее необходимые растению в данный период его жизни.
Для правильного внесения удобрений необходимо знать состав почвы и потребность того или иного вида растений в питательных веществах.
Физиология корня
Рост корня.
Корень обладает неограниченным ростом. Растет он верхушкой, на которой располагается апикальная меристема.
Возьмем 3-4 дневные проростки семян фасоли, нанесем на развивающийся корень тушью тонкие метки на расстоянии 1 мм друг от друга и поместим их во влажную камеру. Через несколько дней можно обнаружить, что расстояние между метками на кончике корня увеличилось, в то время как в более высоко расположенных участках корня оно не меняется. Этот опыт доказывает верхушечный рост корня (рис. 2).
Данный факт используется в практической деятельности человека. При пересадке рассады культурных растений проводят пикировку
- удаление верхушки корня. Эта приводит к прекращению роста главного корня и вызывает усиленное развитие боковых корней (рис 3). В результате всасывающая площадь корневой системы значительно увеличивается, все корни располагаются в верхних наиболее плодородных слоях почвы, что приводит к увеличению урожайности растений.
Поглощение корнем и транспортировка воды и минеральных веществ. Поглощение из почвы и передвижение к наземным органам воды и минеральных веществ - одна из важнейших функций корня. Эта функция возникла у растений в связи с выходом на сушу. Строение корня приспособлено для поглощения воды и элементов питания из почвы. Вода попадает в тело растения через ризодерму, поверхность которой сильно увеличена благодаря наличию корневых волосков. В этой зоне корня формируется проводящая система корня - ксилема, необходимая для обеспечения восходящего тока воды и минеральных веществ.
Поглощение воды и минеральных веществ.
Поглощение воды и минеральных веществ растением происходит независимо друг от друга, так как эти процессы основаны на различных механизмах действия. Вода проходит в клетки корня пассивно, а минеральные вещества поступают в клетки корня в основном в результате активного транспорта, идущего с затратами энергии.
Вода поступает в растение в основном по закону осмоса
. Корневые волоски имеют огромную вакуоль, обладающую большим осмотическим потенциалом,
который обеспечивает поступление воды из почвенного раствора в корневой волосок.
Горизонтальный транспорт веществ.
Поглощению минеральных веществ способствует и выделение корнем различных органических кислот, переводящих неорганические соединения в доступную для поглощения корнями форму.
В корне поперечное движение воды и минеральных веществ осуществляется в следующем порядке: корневой волосок, клетки паренхимы коры, эндодерма, перицикл, паренхима осевого цилиндра, сосуды корня. Горизонтальный транспорт воды и минеральных веществ происходит по трем путям (рис. 4):
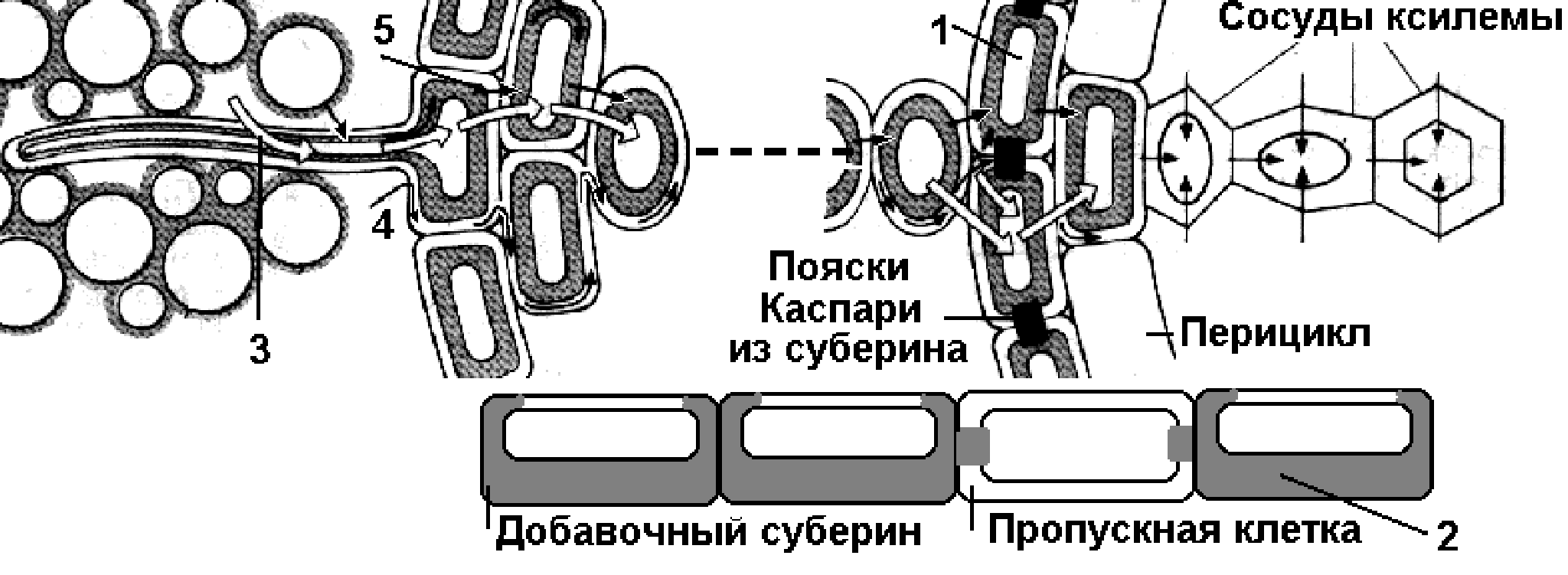
В корне вода передвигается по апопласту до эндодермы. Здесь ее дальнейшему продвижению мешают водонепроницаемые клеточные стенки, пропитанные суберином (пояски Каспари). Поэтому вода попадает в стелу по симпласту через пропускные клетки (вода проходит через плазматическую мембрану под контролем цитоплазмы пропускных клеток эндодермы). Благодаря этому происходит регуляция движения воды и минеральных веществ из почвы в ксилему. В стеле вода уже не встречает сопротивления и поступает в проводящие элементы ксилемы.
Вертикальный транспорт веществ.
Корни не только поглощают воду и минеральные вещества из почвы, но и подают их к надземным органам. Вертикальное перемещение воды происходит по мертвым клеткам, которые не способны толкать воду к листьям. Вертикальный транспорт воды и растворенных веществ обеспечивается деятельностью самого корня и листьев. Корень представляет собой нижний концевой двигатель, подающий воду в сосуды стебля под давлением, называемым корневым. Под корневым давлением
понимают силу, с которой корень нагнетает воду в стебель. Корневое давление возникает главным образом в результате повышения осмотического давления в сосудах корня над осмотическим давлением почвенного раствора. Оно является следствием активного выделения клетками корня минеральных и органических веществ в сосуды. Величина корневого давления обычно - 1-3 атм.
Доказательство наличия корневого давления служит “плач растения” и гуттация.
“Плач растения
” - это выделение жидкости из перерезанного стебля. Такую жидкость называют пасокой
.
Гуттация -
это выделение воды у неповрежденного растения через кончики листьев, когда оно находится во влажной атмосфере или при интенсивном поглощении воды и минеральных веществ из почвы.
Верхний концевой двигатель, обеспечивающий вертикальный транспорт воды - присасывающая сила листьев. Она возникает в результате транспирации
- испарения воды с поверхности листьев. При непрерывном испарении воды создается возможность для нового притока воды к листьям. Сосущая сила листьев у деревьев может достигать 15-20 атм.
В сосудах ксилемы вода движется в виде непрерывных водяных нитей. При движении вверх молекулы воды сцепляются друг с другом (когезия), что заставляет их двигаться друг за другом. Кроме того, молекулы воды способны прилипать к стенкам сосудов (адгезия). Таким образом, поднятие воды по растению осуществляется благодаря верхнему и нижнему двигателям водного тока и силам сцепления молекул воды в сосудах. Основной движущей силой является транспирация.
Запасающие корни. Часто корень выполняет функцию накопления запаса питательных веществ. Такие корни называют запасающими. От типичных корней они отличаются сильным развитием запасающей паренхимы, которая может находиться в первичной (у однодольных) или вторичной коре, а также в древесине или сердцевине (у двудольных). Среди запасающих корней различают корневые клубни и корнеплоды.
- Горизонтальный пунктир показывает границу стебля и корня, черным цветом обозначена ксилема.
орневые клубни характерны как для двудольных, так и для однодольных растений, и образуются в результате видоизменения боковых или придаточных корней (чистяк, ятрышник, любка). Вследствие ограниченного роста в длину они могут иметь овальную, веретеновидную форму и не ветвятся. У большинства видов двудольных и однодольных клубень является лишь частью корня, а на остальном протяжении корень имеет типичное строение и ветвится (батат, георгина, лилейник). - Корнеплод образуется, в основном, в результате утолщения главного корня, но его образовании принимает участие и стебель (рис. 5).
Корнеплоды характерны и для многих культурных овощных, кормовых и технических двулетних растений, и для дикорастущих травянистых многолетних растений (цикорий, одуванчик, скорцонер, женьшень, мак восточный).
Чаще всего корнеплоды образуются в результате вторичного утолщения корней (морковь, пастернак, петрушка, сельдерей, репа, редька, редис). При этом запасающая ткань может развиваться как в ксилеме, так и в флоэме. В утолщении главного корня может принимать участие и перицикл, формируя добавочные камбиальные кольца (у свеклы).
Основные функции корня: обеспечивает закрепление растения в почве, всасывание почвенного водного раствора солей и транспорт его к надземным частям растения.
Дополнительные функции: запасание питательных веществ, фотосинтез, дыхание, вегетативное размножение, выделение, симбиоз с микроорганизмами, грибами. Первые настоящие корни появились у папоротникообразных.
Зародыш корня называется зародышевым корнем и закладывается одновременно с почкой в зародыше семени.
У растений различают:
Главный корень. Он образуется из зародышевого и сохраняется на протяжении всей жизни. Всегда один.
Боковые корни. Ответвляются от корней (главного, дополнительных, боковых). Образуют при ветвлении корни 2-го, 3-го и т. д. порядка.
Придаточные корни. Образуются в любой части растения (стебле, листьях).
Совокупность всех корней растения образует корневую систему. Корневая система формируется в течение всей жизни растения. Ее формирование обеспечивают преимущественно боковые корни. Различают два типа корневой системы: стержневую и мочковатую.
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Меристемы- образовательные ткани- расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля).
Определение размеров корневых систем требует специальных методов. Очень много в этом отношении достигнуто благодаря работам русских физиологов В.Г. Ротмистрова, А.П. Модестова, И.В. Красовской. Оказалось, что общая поверхность корней обычно превышает поверхность надземных органов в 104-150 раз. При выращивании одиночного растения ржи было устоновленно, что общая длинна его корней достигает 600 км., при этом на них образуется 15 млрд. корневых волосков. Эти данные говорят об огромной потенциальной способности к росту корневых систем. Однако эта способность не всегда проявляется. При росте растений в фитоценозах, с достаточно большой густотой их строение, размеры корневых систем заметно уменьшаются.
С физиологической точки зрения корневая система не однородна. Довольно не вся поверхность корня участвует в поглощении волы. В каждом корне различают несколько зон (рис.1). Правда, не всегда все зоны выражены одинаково четко.
Окончание корня с наружи защищено корневым чехликом, напоминающим округлый колпачок, таящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика слущиваются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усилено делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона-зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки, затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а так же расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.
Рис. 1. Схема строения корня:
А - продольный разрез: 1-корневой чехлик; 2- меристема; 3-зона растяжения; 4- зона корневых волосков; 5- зона ветвления;
Б - поперечный разрез (по М.Ф.
Даниловой): 1 - ризодерма; 2 - корневой волосок; 3 - паренхима; 4 - эндодерма; 5-
пояски Каспари; 6 - перицикл; 7 - флоэма; 8 - ксилема. Пунктирные стрелки- пути
передвижения веществ, поглощаемых из наружного раствора. Сплошные стрелки путь
растворов по симпласту; прерывистые - путь по апопласту.
Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системных крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток- эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином. При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения, как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня.. При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление волосков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является основной зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет 5-10 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче.
Корень по длине можно разделить на несколько участков, имеющих различное строение и выполняющих различные функции. Эти участки называют зонами корня. Выделяют корневой чехлик и следующие зоны: деления, растяжения, всасывания и проведения.
Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы конуса нарастания. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует самый поверхностный слой с корневыми волосками - ризодерма (эпиблема). Защитную функцию выполняет ниже лежащая ткань - экзодерма.
В первичном строении корня выделяют три части: ризодерму, первичную кору и
осевой (центральный) цилиндр.
Строение ризодермы рассматривалось в теме "Покровные ткани".
На первичную кору приходится основная масса первичных тканей корня. Ее клетки
накапливают крахмал и другие вещества. Эта ткань содержит многочисленные
межклетники, имеющие значение для аэрации клеток корня. Наружные клетки
первичной коры, лежащие непосредственно под ризодермой, называются экзодермой.
Основная масса коры (мезодерма) образована паренхимными клетками. Самый
внутренний слой носит название эндодермы. Это ряд плотно сомкнутых клеток (без
межклетников).
Центральный или осевой цилиндр (стела) состоит из проводящих тканей, окруженных
одним или несколькими слоями клеток - перициклом.
Внутренняя часть центрального цилиндра у большинства растений занимает сплошной
тяж первичной ксилемы, дающий к перициклу выступы в виде ребер. Между ними
размещаются тяжи первичной флоэмы.
У двудольных и голосеменных растений уже в раннем возрасте в центральном цилиндре корня между ксилемой и флоэмой появляется камбий, деятельность которого приводит к вторичным изменениям и в конечном итоге формируется вторичная структура корня. К центру камбий откладывает клетки вторичной ксилемы, а к периферии - клетки вторичной флоэмы. В результате деятельности камбия первичная флоэма оттесняется кнаружи, а первичная ксилема остается в центре корня.
Вслед за изменениями в центральном цилиндре корня происходят изменения в коровой части. Клетки перицикла начинают делиться по всей окружности, в результате чего возникает слой клеток вторичной меристемы - феллогена (пробкового камбия). Феллоген, в свою очередь, делясь, откладывает наружу феллему, а внутрь - феллодерму. Образуется перидерма, пробковый слой которой изолирует первичную кору от центрального цилиндра. В результате вся первичная кора отмирает и постепенно сбрасывается; наружным слоем корня становится перидерма. Клетки феллодермы и остатки перицикла в дальнейшем разрастаются и составляют паренхимную зону, которую называют вторичной корой корня (рис. 2).
При развитии запасающей паренхимы главного корня происходит формирование запасающих корней или корнеплодов. Различают корнеплоды:
1. Монокамбиальные (редька, морковь) - закладывается только один слой камбия, а запасные вещества могут накапливаться либо в паренхиме ксилемы (ксилемный тип - редька), либо в паренхиме флоэмы (флоэмный тип - морковь);
2. Поликамбиальные - через определенные промежутки времени происходит заложение нового слоя камбия (свекла).
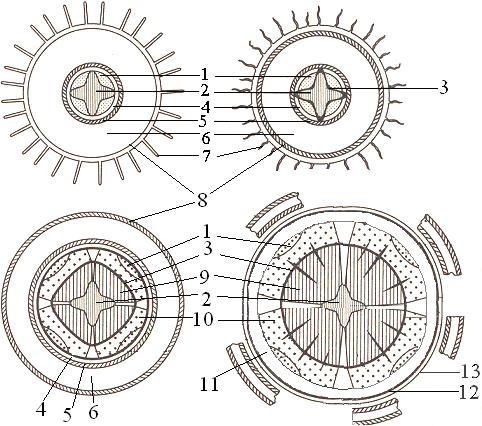
Рис. 2. Переход от первичного строения корня к вторичному:
1 - первичная флоэма, 2 - первичная ксилема, 3 - камбий, 4 - перицикл, 5 - эндодерма, 6 - мезодерма, 7 - ризодерма, 8 - экзодерма, 9 - вторичная ксилема, 10 - вторичная флоэма, 11 - вторичная кора, 12 - феллоген, 13 - феллема.
Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными организмами, в связи с тем что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем.
Большое значение для формирования корневых систем играет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватываются атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков.
Важное значение для развития корневых систем имеет аэрация . Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания . Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений – их усиленному ветвлению.
На продольном разрезе через кончик корня можно обнаружить следующие участки: корневой чехлик , зону ро-ста , зону корневых волосков , зону со строением взрослого корня (рис. 59).
Зона роста расположена несколько отступя от кончика корня. Она составле-на несколькими рядами узких таблитчатых клеток с тонкими оболочками и протоплазмой, заполняющей полость клетки. Это образовательная ткань — меристема . Фактически имеются две меристемы: одна — кончика кор-ня, другая — корневого чехлика.
Клетки последней отчленяют новые клет-ки кнаружи и формируют корневой чехлик . Самые наружные клетки округ-ляются, разъединяются и отшелушиваются под механическим воздействием твердых частиц почвы. Отмершие клетки замещаются клетками, располо-женными ниже их и возникающими из меристемы корневого чехлика. Кнутри из меристемы кончика корня путем деления откладываются клетки собственно корня, из которых по мере дальнейшего развития и роста фор-мируются различные ткани корня. Несколько выше меристемы кончика корня на продольном разрезе уже заметна дифференцировка клеток кор-ня на 3 слоя:
- наружный слой, в дальнейшем развивающийся в кожицу — эпидермис корня;
- средний слой, дающий далее начало прово-дящей части корня;
- промежуточный слой, дальше развивающий так называемую кору корня (рис. 60 в, д, г).
По мере дальнейшего разви-тия эти три зоны внутреннего строения корня хорошо дифференцируются. На поперечном разрезе, проведенном в области развития корневых волосков, корень имеет строение, изображенное на рисунке 61.
Снаружи корень покрыт однослойным эпидермисом . Клетки эпидер-миса живые, имеют тонкие стенки. На периферии они вытянуты в длинные выросты — корневые волоски (рис. 61, а). Ядро клеток перемещается в корневой волосок, располагаясь в переднем, растущем его конце, на ниж-ней стороне (рис. 62).
Корневые волоски имеют длину 0,15—2,5 мм (во влажном воздухе и в искусственной обстановке могут достигать 8 мм). Число их на корне очень велико — в среднем на 1 лш 8 до 425 штук. Зона корневых волосков зани-мает участок корня длиной 1—2 см. Уже на расстоянии 3—8 мм от начала зоны корневые волоски отмирают, отпадают и по мере роста корня ближе к верхушке корня возникают все новые и новые. Корневые волоски выде-ляют наружу различные вещества (кислоты), которые растворяют нераство-римые в воде соединения, имеющиеся в почве, и делают их доступными для растений. Проникая между частицами почвы, корневые волоски плотно срастаются с ними. При выдергивании из земли и при дальнейшей пересад-ке растение часто страдает или даже совсем не приживается вследст-вие отрывания молодых, растущих частей корня с корневыми волос-ками.
За зоной корневых волосков расположена кора корня (рис. 61, к). Она представлена несколькими рядами многогранных паренхимных кле-ток. Самый наружный слой коры корня составлен более крупными, несколь-ко радиально вытянутыми клетками, плотно примыкающими к эпидермису. По опадении корневых волосков оболочки клеток этого слоя опробкове-вают; кроме того, под ними возникает слой пробковых клеток , которые, таким образом, защищают наружный слой корня от испарения. Глубоко лежащие слои корня состоят из более мелких клеток.
Самый внутренний слой коры образует так называемую эндодерму , отчленяющую кору от срединной части корня, так называемого центрального цилиндра , или стели (рис. 61, эн). Клетки эндо-дермы расположены в один слой и тесно придвинуты друг к другу. На-ружные стенки их тонкие, внутренние же и радиальные (боковые) стенки получают сильное утолщение и, пропитываясь суберином, опробковева-ют, становясь непроницаемыми для воды и газов. Среди клеток эндодермы встречаются отдельные клетки с неутолщенными стенками, богатые содер-жимым (пропускные клетки ); через эти клетки происходит про-никновение воды из коровой части корня в центральный цилиндр.
Центральный цилиндр, занимая срединную часть корня, состоит из различных тканей. Самый наруж-ный слой (реже два слоя) клеток, примыкающий изнутри к эндодер-ме, носит название перицикла (рис. 61). Клетки его более крупные, слегка вытянуты по радиусу и имеют тонкие клетчатковые оболочки. В этом слое про-исходит заложение боковых ко-решков, почему перицикл часто называют корнеродным слоем . Образуются они следую-щим образом: над группой сосу-дов в перицикле наблюдается уси-ленное деление его клеток, бла-годаря чему образуется небольшой бугорок, выпяченный в перифери-ческую сторону корня. Снаружи его облегают клетки эндодермы.
Постепенно в бугорке формируются слои, свойственные меристеме глав-ного корня, из которых в дальнейшем образуются соответственные ткани. Бугорок продолжает расти, окруженный слоем эндодермы (так называемый «кармашек»), и, прорывая кору, выходит наружу. Материал с сайта
Таким образом, боковые корни, в отличие от главного корня и стебля, возникают эндогенно (внутриродно). Эндогенно возникают все при-даточные корни, из какой бы части растения они ни образовались.
За перициклом расположена паренхимная ткань, в которую погружены проводящие элементы (сосудистые пучки ). Сосудистый пучок корня построен по радиальному типу (радиальный пучок). Ксилема его расположена группами, лучами, идущими от периферии к центру. Число лучей обычно небольшое — 3, 4, 5. Иногда группы ксилемы не доходят до центра, который заполнен тогда паренхимными клетками. Сосуды ксилемных лучей имеют более узкие просветы ближе к периферии и более крупные к центру. Наружные сосуды закладываются раньше цен-тральных. Развитие ксилемы в корне идет от периферии к центру. Наружные сосуды обычно бывают кольчатыми или спиральными; более крупные вну-тренние — пористые. Между лучами ксилемы в верхней части расположе-ны участки флоэмы, составленной ситовидными трубками с сопровождаю-щими клетками и лубяной паренхимой.
На этой странице материал по темам:


